Le maïs cireux est un maïs hybride dont l'amidon du grain est entièrement constitué d'amylopectine et qui présente un intérêt particulier pour certaines préparations industrielles.
Ce maïs est généralement placé dans le Zea mays Ceratina Group. Il a été décrit comme convariété ou variété de Zea mays (Zea mays var. ceratina Kuleshov) mais n'est généralement plus accepté comme tel
Le maïs denté traditionnel contient en moyenne 25% d'amylose et 75% d'amylopectine, et c'est cette dernière qui donne la gluance recherchée par les amidonniers. Or il est très difficile et coûteux de séparer ces deux substances, d'où l'idée d'utiliser le maïs cireux qui produit 100% d'amylopectine.
L'amylopectine est utilisée dans les produits alimentaires, textiles, adhésifs ainsi que dans l'industrie du papier et du carton ondulé. Peu après la guerre, des essais montrèrent que l'affourragement à base de maïs cireux permet un engraissement plus rapide que le maïs denté ne le permet. Le maïs cireux connut alors un vif gain d'intérêt.
Les généticiens purent démontrer que ce maïs a une déficience dans le métabolisme empêchant la synthèse d'amylose dans l'albumen. Elle est codée par un unique gène (wx). Le maïs cireux donne un rendement de 3,5 % inférieur à ses congénères dentés et se doit d'être planté à au moins deux cents mètres d'un champ de maïs conventionnel si on veut éviter l'hybridation.
Histoire
Le maïs cireux a été découvert en Chine en 1909. Comme cette plante possédait différentes particularités, les sélectionneurs américains l'utilisèrent longtemps comme marqueur génétique afin de rendre visible certains gènes cachés dans d'autres programmes de sélection du maïs. En 1922, un chercheur trouva que l'albumen du maïs cireux ne contenait que de l'amylopectine et pas d'amylose contrairement aux variétés conventionnelles de maïs qui contiennent les deux. Jusqu'à la Seconde Guerre mondiale la principale source d'amylopectine était le manioc mais lorsque les Japonais coupèrent les lignes d'approvisionnement des États-Unis, l'industrie américaine dut se tourner vers le maïs cireux.
Le maïs cireux se trouve mentionné pour la première fois dans les archives du ministère américain de l'agriculture (USDA), mais l'histoire exacte de cette variété de maïs est inconnue.
En 1908, le père J. M. W. Farnham, missionnaire presbytérien à Shanghai, envoya un échantillon de graines à l' U.S. Office of Foreign Seed and Plant Introduction. Une note annexée le présentait ainsi :
« Type particulier de maïs. Il y a plusieurs couleurs, mais on dit qu'elles sont toutes de la même variété. Le grain est, me semble-t-il, beaucoup plus visqueux que dans les autres variétés et peut convenir à certains usages, peut-être en bouillie. ».
Ces graines ont été semées le 9 mai 1908, près de Washington, D.C., par un botaniste du nom de G.N. Collins. Il réussit à mener 53 plants à maturité et en fit une description détaillée, accompagnée de photographies, qui fut publiée dans un bulletin de l'USDA paru en décembre 1909. En 1915, la plante fut redécouverte en Haute-Birmanie et à nouveau en 1920 aux Philippines. Kuleshov, lorsqu'il vérifia la distribution du maïs en Asie, la trouva en de nombreux autres endroits.
La découverte en Chine d'un type de maïs distinct posa la question historique de savoir si le maïs était connu en Extrême-Orient avant la découverte de l'Amérique. La controverse fut close à la fin du XIX siècle par De Candolle qui déclara que : « le maïs est d'origine américaine, et n'a été introduit dans l'ancien monde que depuis la découverte du nouveau. Je considère ces deux assertions comme positives, malgré l'opinion contraire de quelques auteurs ». Mais la découverte de cette variété unique de maïs suggéra un ré-examen de la question. Il indiqua également que les Portugais sont arrivés en Chine en 1516, y introduisant simultanément le maïs.
Collins a émis l'hypothèse que le maïs cireux est apparu à la suite d'une mutation en Haute-Birmanie. Dans cette hypothèse, il semblait difficile de concevoir que depuis 1516 cette plante américaine ait eu le temps de pénétrer dans un pays sauvage inaccessible aux étrangers, de produire une mutation, et ensuite, en tant que mutant de se répandre depuis les Philippines jusqu'au nord de la Mandchourie et à la région de Primorsky en l'espace de trois ou quatre centaines d'années. On peut aujourd'hui répondre à ces deux arguments. D'abord, on sait que la mutation waxy est très commune (voir paragraphe #Génétique). Ensuite, le fait que le maïs, même introduit en Asie après l'époque de Christophe Colomb, a dû être rapidement accepté par les populations arriérées indique seulement que, à l'instar de la pomme de terre en Irlande, il a rencontré un besoin aigu et pressant. Selon Goodrich il existe en Chine quelque 6000 histoires locales écrites depuis 347. La première description précise du maïs dans ces publications date du XVI siècle.
Ho, éminent historien chinois, indiqua : « en résumé de l'introduction du maïs en Chine, nous pouvons dire que le maïs fut introduit en Chine deux ou trois décennies avant 1550... » Il se pourrait que, comme l'affirment certaines études, le maïs soit arrivé en Asie avant 1492, mais à l'heure actuelle pas un seul fragment de plante, ni aucun objet, aucune illustration ou trace écrite, n'a été porté à notre connaissance pour le prouver. Par conséquent, toute affirmation non documentée que cet événement ce serait produit à une époque antérieure doit être considérée avec scepticisme jusqu'à ce qu'elle soit prouvée. Ainsi, les deux assertions de De Candolle restent valides.
Dans sa publication, Collins a caractérisé cette nouvelle plante par la possession de nombreux caractères uniques. Aucune trace de ces caractères dans les formes enregistrées de Zea mays n'avait jusqu'alors été trouvée. La combinaison de plusieurs de ces caractères confère à la plante une résistance au dessèchement des soies par les vents secs et chauds au moment de la floraison. Bien que ces plantes produisent de si petits épis qu'elles ne pouvaient pas entrer en compétition directe avec les variétés améliorées, la présence de cette adaptation leur donnait un intérêt économique, particulièrement dans certaines régions semi-arides du Sud-Ouest des États-Unis. On a donc cherché à introduire par hybridation les caractères désirables de cette petite variété dans les types de maïs plus grands et plus productifs.
Quand Collins découvrit une telle différence d'aspect de l'albumen entre un maïs cireux et un type normal, il soupçonna une différence dans la composition chimique, mais les analyses ne montraient aucun résultat inhabituel. Les pourcentages d'amidon, d'huile et de protéines étaient tous dans les normes habituelles. Pourtant la nature physique de l'amidon l'intriguait et il écrivit : « Au vu du récent développement de produits de maïs spécialisés pour l'alimentation humaine, le type unique d'amidon peut avoir une certaine importance économique ».
En fait, pendant de nombreuses années la principale utilisation du maïs cireux a été celle de marqueur génétique pour d'autres programmes de sélection de maïs. Les sélectionneurs étaient en mesure d'utiliser certains caractères pour « étiqueter » l'existence de gènes cachés et les suivre dans les programmes de sélection. Le maïs cireux aurait pu disparaïre à nouveau aux États-Unis sans cette application particulière dans la sélection.
En 1922, un autre chercheur, P. Weatherwax de l'université de l'Indiana à Bloomington, signala que l'amidon du maïs cireux était entièrement d'une forme « rare » appelée « érythrodextrine », connue aujourd'hui sous le nom d'amylopectine. Il découvrit que cet amidon rare se teintait en rouge en présence d'iode, alors que l'amidon normal se teinte en bleu. Bates, French et al. et Sprague, Brimhall, et al. confirmèrent que l'amidon de l'endosperme du maïs cireux est constitué presque exclusivement d'amylopectine. La présence de l'amylopectine dans le riz avait été montrée précédemment par Parnell.
En 1937, juste avant la Seconde Guerre mondiale, G.F. Sprague et d'autres sélectionneurs de ce qui était alors le Iowa State College avait entrepris un programme de croisements pour essayer d'introduire le caractère cireux dans un maïs hybride à haut rendement. À cette époque, le type cireux n'avait plus les traits structurels particuliers notés par Collins, probablement à cause des années d'hybridations avec des stocks génétiques variés. Seul, l'endosperme unique avait été conservé.
Le maïs cireux n'avait pas alors une très grande importance car la source principale d'amylopectine pure était encore le manioc, arbuste tropical à gros tubercule souterrain. Cette situation perdura jusqu'à la Seconde Guerre mondiale, quand les Japonais coupèrent les lignes d'approvisionnement des États-Unis et forcèrent les transformateurs à se tourner vers le maïs cireux. Ce dernier parut alors particulièrement adapté à cet objectif parce qu'il pouvait être moulu avec les mêmes équipements déjà largement utilisés pour le maïs ordinaire.
Selon H. H. Schopmeyer la production de maïs cireux dans l'Iowa à des fins industrielles s'élevait à 356 tonnes en 1942 et 2 540 tonnes en 1943. En 1944, il existait seulement cinq variétés de maïs cireux aptes à la production d'amidon cireux. En 1943, pour couvrir tous les besoins en amylopectine, environ 81 650 tonnes de grains furent produites. Depuis la Seconde Guerre mondiale jusqu'en 1971, tout le maïs cireux produit aux États-Unis l'était dans le cadre de contrats avec des transformateurs pour l'industrie ou le secteur alimentaire. En fait, l'essentiel de ce maïs était cultivé dans un petit nombre de comtés des États de l'Iowa, de l'Illinois et de l'Indiana. Mais en 1970, une épidémie d'helminthosporiose du maïs (Helminthosporium maydis Nisik. and Miyake) sévit dans toute la Corn Belt américaine.
Au même moment, au moins 80 % du maïs cultivé aux États-Unis était sensible à l'helminthosporiose parce que ce maïs contenait un cytoplasme mâle-stérile du « type Texas », qui permettait la production de semences hybrides sans qu'il soit nécessaire de procéder à la castration mécanique ou manuelle. Aussi tout naturellement, il y eut un véritable affollement en 1971 pour trouver un type de maïs au cytoplasme normal – cytoplasme résistant à l'helminthosporiose. C'est ainsi que des semences de maïs cireux sont entrées dans le marché.
Des rétrocroisements ont aussi été réalisés largement pour transférer des gènes particuliers tels que wx (waxy), o2 et le gène Ht de résistance à l'helminthopsoriose. Certains agriculteurs qui ont alimenté leur bétail bovin avec des grains de maïs cireux ont remarqué que les animaux profitaient mieux. Des essais d'alimentation furent organisés qui démontrèrent que le maïs cireux entraînait de meilleurs gains de poids que le maïs normal denté. L'intérêt pour le maïs cireux explosa subitement, et ce type de maïs perdit son statut de curiosité botanique et de production spécialisée pour devenir l'objet de recherches de la plus grande importance. En 2002, une production estimée entre 1,2 et 1,3 million de tonnes fut réalisée aux États-Unis sur environ 2 000 km, représentant seulement 0,5 % de la production totale de maïs américaine.
Biologie
La composition des maïs varient selon les cultivars et en fonction du développement du grain. Par exemple, le pourcentage d'amylose va de 20 à 36 % pour 399 cultivars de maïs normal. Il existe des sources de plasma germinatif de maïs qui vont de moins de 20 % à 100 % de complément d'amylopectine.
L'endosperme cireux est en soi un défaut du métabolisme, et sa faible fréquence chez la plupart des populations de maïs face à des mutations récurrentes indique qu'il s'exprime contre la sélection naturelle.
Maïs chinois
Collins a noté, entre autres, les caractères suivants, inhabituels, du maïs chinois :
plusieurs traits structurels uniques qui permettent aux plantes de résister au desséchement des soies par le vent au stade de la floraison ;
un port particulier, en ce sens que les quatre ou cinq feuilles supérieures apparaissent toutes du même côté de la tige principale de la plante ; les feuilles des nœuds supérieurs sont très érigées, alors que les feuilles basses sont plus étalées et retombantes ;
l'une des principales choses qu'il a noté est la composition de l'albumen des grains de maïs ; à ce sujet il écrivit :
« La texture de l'albumen est l'un des traits uniques de ce maïs. Coupé selon n'importe quelle direction, il se sépare en suivant une sorte de clivage, exposant une surface terne et lisse ; la texture évoque celle des cires les plus dures, bien qu'elle soit encore plus dure et plus cristalline. C'est cette ressemblance optique qui suggéra le terme d'albumen cireux. »
Le taux d'humidité du grain doit être au plus égal à 16 % pour que le caractère cireux puisse être reconnu visuellement.
Dérive génétique
Des expériences menées par Sprague ont montré que dix à vingt plantes sont nécessaires pour disposer d'une représentation adéquate de la diversité génétique dans une variété de maïs à pollinisation ouverte. Comme le nombre d'épis conservés comme semence par les anciens cultivateurs asiatiques qui ne disposaient que de petits carrés de terre était souvent inférieure et, en fait, vu que de nouvelles populations de maïs sont parfois créées en semant la descendance d'un seul épi, il s'ensuit qu'il dut se produire souvent une dérive génétique dans la fréquence des gènes résultant de la création de petits peuplements de sélection.
Un exemple frappant de dérive génétique chez le maïs est l'occurrence dans certaines parties de l'Asie de variétés à endosperme cireux. De telles variétés sont inconnues chez les maïs américains, mais le caractère cireux lui a été découvert chez des variétés non cireuses : chez un maïs flint de Nouvelle-Angleterre et chez une variété sud-américaine.
Le fait que le maïs cireux se trouve si communément dans une partie du monde qui possède aussi des variétés cireuses de riz, de sorgho et de millet peut être attribué à une sélection artificielle. Les habitants de l'Asie étant familiers des variétés cireuses de ces céréales et accoutumés à les utiliser pour des usages particuliers ont reconnu le caractère cireux du maïs à la suite de son introduction en Asie après la découverte de l'Amérique et ont délibérément isolé des variétés purement pour leur endosperme cireux. Mais le fait que l'endosperme cireux ait attiré leur attention la première fois est probablement dû à la dérive génétique. Le gène de l'endosperme cireux, qui a une fréquence faible chez les maïs américains, a apparemment atteint une fréquence élevée dans certains échantillons de maïs asiatiques.
En fait, la pratique rapportée par Stonor and Anderson de cultiver le maïs sous forme de plantes isolées parmi d'autres céréales dut entraîner une certaine part d'autopollinisation et, dans tout stock dans lequel le gène cireux était présent, devait inévitablement conduire en un très court laps de temps à l'établissement de variétés pures de maïs cireux aux propriétés spéciales que des gens habitués aux caractères cireux d'autres céréales pouvaient difficilement manquer de reconnaître
Génétique
Un unique gène récessif waxy (wx), situé sur le bras le plus court du chromosome 9, code l'albumen cireux du grain de maïs (l'allèle Wx code un albumen à amidon normal). Ce fait a été démontré en premier par Collins et Kempton. La structure du locus du type cireux sauvage (wx+) a été déterminée par analyse séquentielle de l'ADN. Le gène a 3718 paires de bases (14 exons et 13 introns). L'endosperme cireux est l'équivalent pour le maïs du caractère « gluant » chez le riz. De nombreuses autres espèces présentent aussi cette mutation « cireuse », parmi lesquelles, outre le riz, le sorgho, le millet, l'orge et le blé. Elle est toujours caractérisée par la coloration bleue des granules d'amidon en présence d'iode.
Lors de croisements entre des plantes hétérozygotes pour le caractère cireux, on constate un petit, mais significatif, écart par rapport au ratio mendélien attendu en auto-pollinisation Bear obtint, à partir de la 71 épis ségrégués de la génération F1, 23,77 % de grains cireux et 76,23 % grains non cireux. Cela est mis en évidence par les deux types hétérozygotes, Wx Wx wx et wx wx Wx. Le gène waxy est épistatique pour tous les autres gènes mutants connus formant de l'amidon tels que dull (du), sugary-1 (su1 ) et sugary-2 (su2), il augmente les sucres et les polysaccharides solubles dans l'eau avec su1 et il provoque une augmentation spectaculaire des sucres et une réduction de l'amidon avec ae ou ae du. La mutation de Wx à wx n'est pas rare chez les variétés cultivées dans la Corn Belt, Bear ayant trouvé trois mutations séparées en wx en trois années consécutives dans une population totale de quelque 100 000 épis.
Mangelsdorf découvrit également de nombreux mutants dans ses champs d'essais. L'allèle « cireux d'Argentine » (wx-a) du locus waxy signalé d'abord par Andrés et Bascialli, est connu pour produire une petite quantité d'amylose (< 5 %) et donne une coloration intermédiaire avec l'iode. D'autres allèles mutants du locus waxy ont été signalés, qui possèdent des amidons aux propriétés similaires à celles observées avec l'allèle wx. On connait plus de 40 allèles mutants pour le locus waxy, ce qui en fait le plus bel ensemble de mutations trouvé chez les plantes supérieures. Certains d'entre eux sont très stables alors d'autres sont très instables.
Le phénotype des mutants stables reste inchangé tandis que celui des mutants instables change du fait de l'insertion d'éléments transposables (5-8). Pour connaître la liste de toutes ces mutations, l'excellent livre de Neuffer, Coe et al. est hautement recommandé. Du fait que la mutation waxy s'exprime par un phénotype non létal et facilement identifiable, elle a fait l'objet d'importantes recherches au cours du XXe siècle. Nelson a réalisé une carte détaillée de la structure génétique de la plupart de ces mutations.
Génotype et caractérisation à l'iode
Le locus wx s'exprime dans l'endosperme, dans le gamétophyte mâle (pollen) ainsi que dans le gamétophyte femelle (sac embryonnaire). L'amylose et l'amylopectine ont des propriétés de fixation de l'iode différentes, l'amylose et l'amylopectine de maïs donnant des valeurs d'affinité pour l'iode d'environ 19 à 20 et 1 %, respectivement. Weatherwax découvrit cette réaction en 1922.
La quantité d'amylose apparente peut être déterminée soit en mesurant l'absorbance du complexe amidon-iode et en rapportant cette valeur à la référence de l'amylose ou l'amylopectine pures ou en mesurant la quantité d'iode (mg) fixée pour 100 mg d'amidon par un titrage potentiométrique et en comparant la valeur trouvée à la quantité fixée par une amylose standard. Les valeurs utilisées pour la fixation de l'iode, cependant, sont seulement des estimations du contenu en amylose du fait des différences dans la capacité de fixation (et la structure) de l'amylose et de l'amylopectine selon les types d'amidon. Par exemple, les molécules d'amylopectine à longues branches externes fixent plus d'iode que celles à branches courtes, d'où une faible mesure de l'amylose apparente. Les profils chromatographiques d'amidons de type wx, toutefois, ne révèlent aucun pic d'amylose.
Les plants hétérozygotes pour le gène waxy (Wx:wx) peuvent être caractérisés en traitant le pollen à l'iode. La moitié du pollen sera bleue et l'autre moitié brune tandis que les grains resteront bleus (très utile dans les programmes de rétrocroisement). Si la plante est homozygote récessive (wx:wx), tout le pollen sera brun et les grains aussi. Chez les homozygotes dominants (Wx:Wx), l'iode deviendra bleue.
Aspects agronomiques
La culture du maïs dont l'amidon est constitué d'amylopectine pure n'est pas simple. Le gène waxy étant récessif, le maïs cireux doit être isolé d'au minimum 200 mètres de tout champ de maïs normal. Cependant, dans la monoculture du maïs, les semis naturels ne sont pas rares dans les régions sans gel, or quelques repousses dans un champ de maïs cireux suffisent pour contaminer tout le champ même s'il est isolé. Une solution pourrait être de faire des 15 à 20 rangées extérieures du champ une bande tampon, qui peut servir à nourrir le bétail. La nécessité de détecter toute contamination par le maïs normal justifierait la création de variétés cireuses à grains blancs, ce qui permettrait le tri des grains par des cellules photoélectriques et le rejet des semences contaminées. Mais cette idée n'a pas eu d'application connue.
La réponse aux fertilisants, aux maladies, aux insectes ravageurs, aux stress environnementaux, etc., ne devrait en théorie marquer aucune différence entre les hybrides de maïs cireux et ses équivalents dentés du fait qu'aucune différence n'existe entre ces deux types de maïs, sauf dans le grain. Le sucrose est le sucre de transport, mais il est normalement converti en amidon lorsqu'il est transféré dans l'endosperme du maïs.
Utilisation
L'amylopectine : usages industriels
L'amidon est le principal hydrate de carbone de réserve dans le règne végétal. Bien que cette substance soit présente chez la plupart des espèces végétales, seules quelques-unes sont utilisées pour la production commerciale d'amidon, et le maïs est la principale source de l'amidon produit industriellement dans le monde. Au second rang, se trouve la pomme de terre, suivie du blé et, dans une moindre mesure, du riz. L'amidon de maïs a été produit pour la première fois aux États-Unis en 1844 dans l'usine de William Colgate à Jersey City (New Jersey).
Le rendement en amylopectine du grain de maïs va de 58,5 à 69 % (de la matière sèche). La mouture humide de maïs cireux donne des rendements de seulement 90 % par rapport à ceux du maïs denté. L'amidon wx est relativement facile à gélatiniser et produit une pâte visqueuse claire à la surface collante et poisseuse qui ressemble à celles produites par les amidons de racines ou tubercules, telles que pomme de terre et tapioca. La plupart des amidons dans leur forme naturelle ou non modifiée ont des emplois limités dans les diverses industries. La plupart des amidons dont les amidons de maïs cireux sont modifiés soit pour améliorer leurs propriétés intrinsèques, soit pour les utiliser, en fonction des nécessités requises par les applications particulières. De nombreux types d'amidons cireux modifiés ont une multitude d'applications dans les industries du papier, du textile, du carton ondulé, et des adhésifs sans compter l'énorme gamme des applications dans l'agro-alimentaire.
Produits alimentaires
Les amidons modifiés de maïs cireux ont d'importantes applications dans le domaine alimentaire, notamment l'amélioration de l'uniformité, de la stabilité et de la texture de divers produits alimentaires. La limpidité et la stabilité de l'amidon d'amylopectine en fait un produit particulièrement adapté comme épaississeur pour les tartes aux fruits. Il améliore l'onctuosité et le crémeux des conserves et des produits laitiers ainsi que la résistance à la congélation-décongélation des produits surgelés. Il donne une texture et un aspect plus agréables aux aliments secs et aux mélanges. L'amidon de maïs cireux est aussi la matière première préférée pour la production de maltodextrines pour sa meilleure solubilité dans l'eau après déshydratation et sa plus grande stabilité et limpididité en solution.
Industrie textile
L'amidon cireux présente un intérêt pour l'industrie textile du fait de sa capacité à produire des films transparents [38].6.1.3 L'amidon de maïs cireux pour l'industrie des adhésifs diffère de l'amidon de maïs habituel à la fois par sa structure moléculaire et par ses caractéristiques d'empâtage. Selon Watson [67], les pâtes faites à base d'amidon cireux sont longues et cohésives, alors que celles d'amidon de maïs normal sont courtes et épaisses. L'amidon de maïs cireux est l'amidon principal des adhésifs utilisés pour les étiquettes de bouteilles. Ces adhésifs à base d'amidon cireux donnent aux étiquettes une résistance à la resolubilisation qui prévient leur décollage de la bouteille si elles sont mouillées ou soumises à une forte humidité. De plus, les amidons de maïs cireux sont couramment utilisés pour la fabrication de bandes gommées et d'adhésifs pour enveloppes.
Papeterie
Les principaux progrès dans la technologie de la fabrication du papier sont en partie liés à la disponibilité de nouveaux types d'amidons modifiés de maïs cireux. Ces amidons apportent essentiellement des qualités de liant au processus de fabrication du papier, mais ils ajoutent aussi d'autres caractéristiques essentielles, notamment l'amélioration de la force du papier et de ses propriétés d'impression [42].
Amidons
Bien qu'il existe déjà une large gamme d'amidons modifiés de maïs normal ou cireux, il y a un intérêt croissant pour la production d'amidons entièrement naturels. En fait, les consommateurs les préfèrent et les règles d'approbation des processus chimiques sont de plus en plus contraignantes, ce qui encourage la recherche de matériaux d'origine naturelle pour remplacer partiellement ceux chimiquement modifiés. Plusieurs brevets ont été déposés sur des doubles mutants de mais. Deux ont été déposés sur le génotype de maïs wxsu2, l'un pour utiliser l'amidon comme épaississant avec une stabilité à basse température améliorée, et l'autre contre le rassissement du pain. Dans le second cas, le pain a une mie plus moelleuse, humide après la cuisson et une texture et un aspect plus frais après conservation que le pain produit sans cet additif. Plusieurs brevets de la société American Maize-products Company sont basés sur de nouveaux amidons qui peuvent remplacer ceux chimiquement modifiés, contenant tous le gène wx [25–28]. Ces amidons présentent tous une modification de la viscosité, de la limpidité, de l'aspect de la pâte et de la résistance au cycle congélation-décongélation.
Alimentation animale
L'alimentation des animaux à base de maïs cireux a débuté aux États-Unis dans les années 1940. Un rapport de recherches paru en 1944 indiquait que le maïs cireux semblait avoir le pouvoir d'améliorer l'efficacité de la conversion alimentaire. De nombreux autres essais alimentaires impliquant les porcins, les bovins à viande ou à lait ainsi que les veaux et la volaille ont été conçus pour comparer la valeur alimentaire du maïs cireux à celle du grain denté normal [11;32;46;54]. En général, ils ont démontré un avantage pour le maïs cireux. Rarement les investigations ont montré des effets négatifs ou contraires de l'alimentation avec des grains de maïs cireux. Des augmentations à la fois de la production de lait et de la teneur butyrique ne sont pas rares quand le maïs cireux est donné aux vaches laitières en période de lactation. Des augmentations de plus de 20 % du gain de poids moyen quotidien dans l'engraissement des veaux ont été observés lorsque le maïs cireux était comparé au maïs denté normal. En outre, une augmentation de 14 % de l'efficacité alimentaire a été notée en faveur du maïs cireux.
 词典释义:
词典释义:

 ,
,  质
质

 ,
,  黄色
黄色
 黄
黄 面
面
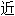
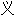 词:
词: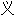 词:
词: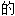 ,浅黄
,浅黄 ,带黄色
,带黄色 ;
; ,
, 白色
白色 ;
; ,淡红色
,淡红色 ,浅红色
,浅红色 ;
; ,暗绿色
,暗绿色 ;
; ;
; ,黯淡
,黯淡 ,无光泽
,无光泽 ;
; ;
; ;
; ,棕色
,棕色 ,栗色
,栗色 ;
; ,苍白
,苍白 ;
; 样
样 ,
,  状
状 ;
;  质
质 ; 产
; 产
 样管型
样管型