La tige est chez les plantes, l'axe généralement aérien ou souterrain (tubercule caulinaire, rhizome), qui prolonge la racine et porte les bourgeons et les feuilles. La tige se ramifie généralement en branches et rameaux formant l'appareil caulinaire. Chez les arbres et les plantes ligneuses on distingue le tronc, partie principale généralement dénudée à sa base, du houppier, formé de l'ensemble branches maîtresses et rameaux. Une plante dont la tige est absente ou très réduite est dite acaule.
La tige diffère de la racine par la présence de nœuds où s'insèrent les bourgeons axillaires et les feuilles, par l'absence de coiffe terminale et par sa structure anatomique. La transition entre racine et tige se fait dans le « collet ». Il peut exister des tiges souterraines comme il existe des racines aériennes.
Par son mode de croissance et de ramification, la tige détermine le port de la plante ; elle assure une fonction de soutien et une fonction de transport des éléments nutritifs entre les racines et les feuilles.
Les tiges creuses sont un habitat pour de nombreuses espèces qui y hibernent ou y pondent leurs œufs. Ces espèces sont dites caulicoles.
Structure anatomique
Coupe transversale d'une tige de lin. 1-moelle 2-protoxylème 3-xylème primaire 4-phloème primaire 5-fibres de sclérenchyme 6-écorce 7-épiderme
On peut observer au microscope l'anatomie des tiges grâce à des coupes transversales fines de tiges ou de pédoncules floraux. Ces coupes peuvent être colorées pour permettre de différencier les cellules et leurs constituants, le plus souvent grâce à la double coloration carmin-vert d'iode (colorant de Mirande).
Ce colorant colore en rose les structures cellulosiques qui sont des tissus vivants (comme les parenchymes, le phloème(4) primaire et secondaire et les collenchymes) et en vert les structures contenant des molécules apparentées aux lipides (tissus subérifiés ou lignifiés comme les parenchymes lignifiés, les sclérenchymes, le xylème(2,3) primaire et secondaire) et en jaunâtre le suber (appelé aussi liège).
La tige est un organe cylindrique à symétrie axiale généralement, sauf dans le cas des tiges carrés ou triangulaires où la symétrie est dite bilatérale (ex : Allium triquetrum)
Les tiges peuvent comporter uniquement des structures primaires (cas des monocotylédones et des dicotylédones jeunes) ou aussi des structures secondaires (dicotylédones âgées). L'organisation de ces structures évite le flambage de la tige par le phénomène de tenségrité. La présence de collenchyme apporte une grande résistance à la flexion et traction (caractéristique chez la menthe, la sauge).
Structure primaire
Une tige comporte 2 zones principales : l'écorce(6) (ou cortex ou encore cylindre périphérique) et le cylindre central occupant, entre autres, les rôles respectifs de protection et de sustentation.
Écorce
L'écorce est réduite dans les tiges, dans lesquelles le cylindre central est très développé, contrairement aux racines où c'est le contraire.
L'écorce est constituée d'un tissu de revêtement d'origine primaire : l'épiderme(7). Ce dernier se compose d'une seule assise (couche) de cellules alignées côte à côte ; on peut remarquer des stomates de temps en temps, ils permettent des échanges gazeux entre le milieu extérieur et la tige.
Sous l'épiderme se trouve le parenchyme cortical, un tissu composé de cellules plus grosses et moins bien organisées. Il est le siège de la photosynthèse et peut servir de réserve à la plante. Souvent on trouve dessous un parenchyme lignifié (il apparait plus vert sur la coupe) qui sert à soutenir la plante, c'est le sclérenchyme(5). Dans les organes jeunes, on trouve le collenchyme.
Cylindre central
La limite entre le cortex et le cylindre central est constituée par les faisceaux conducteurs. Ces derniers se trouvent dans un autre parenchyme appelé parenchyme médullaire. Les faisceaux sont composés de deux types de vaisseaux : le xylème(2,3) et le phloème(4). Le xylème conduit la sève brute composée d'eau, de sels minéraux et de quelques acides aminés des racines vers les organes photosynthétiques où cette sève se charge en sucre. Elle se transforme alors en sève élaborée qui est conduite par le phloème vers les organes demandeurs en énergie tels que les bourgeons, les racines, les tubercules, les fleurs, les fruits... Dans les tiges, le xylème et le phloème sont superposés (le phloème se trouve au-dessus du xylème), contrairement aux racines où ils sont disposés l'un à côté de l'autre. Chez les monocotylédones, ces vaisseaux sont nombreux, disposés sur plusieurs cercles, de taille variable et plus petite. Chez les dicotylédones, le nombre de faisceaux vasculaires est moindre (moins de 8) et ils sont tous sur un même cercle.
Au centre de la tige se trouve la moelle(1) ou zone médullaire qui contient des parenchymes de réserve. Chez la plupart des Poacées (graminées), cette zone est remplacée par une lacune centrale et on appelle alors la tige « chaume ». Cela explique pourquoi la tige du blé est creuse.
Structure secondaire
Les structures secondaires qui peuvent se développer chez un végétal sont de deux sortes : les tissus secondaires de conduction ou les tissus secondaires de revêtement.
Les tissus secondaires de conductions se mettent en place à partir d'un méristème secondaire : le cambium. Il donnera le Xylème secondaire et le Phloème secondaire.
Les tissus secondaires de revêtement sont produits par le phellogène, qui donnera le phelloderme vers l’intérieur et du suber vers l’extérieur.
Croissance et ramification
Croissance primaire (en longueur)
La croissance des entre-nœuds se fait sous l'effet des gibbérellines.
Croissance radiale

Croissance radiale, platane
La croissance secondaire (radiale) est,
Inexistante chez les plantes appartenant à la classe des Liliopsida (Monocotylédones angiospermes)
Ex. de familles : Poaceae (graminées), Arecaceae (palmiers), Cyperaceae (papyrus), Musaceae (bananiers), ...
Les plantes appartenant à cette classe ne possèdent pas de cambium et ne produisent donc ni bois ni racines secondaires (racine poussant perpendiculairement sur une autre racine). La seule façon pour ces plantes à fleurs de pouvoir augmenter leur assise et la rigidité de leur tige, est d'accumuler des tissus morts autour de celle-ci. (comme chez les palmiers)
Importante chez les plantes appartenant à la classe des Magnoliopsida (Dicotylédones angiospermes)
Ex. de familles : Fagaceae (chênes, hêtres, châtaigniers), Cactaceae (cactus), Salicaceae (peupliers, saules), Oleaceae (frênes, oliviers, lilas), Rosaceae (rosiers, pruniers, pommiers, poiriers, cerisiers, ronciers, framboisiers), ...
Les plantes appartenant à cette classe forment un cordon de cellules peu différenciées et non épaissies au sein du faisceau libéro-ligneux, entre le phloème et le xylème appelé procambium. Le procambium, en se divisant, met en place un cambium intrafasciculaire qui formera du xylème secondaire vers l'intérieur et du phloème secondaire vers l'extérieur. L'activité méristèmatique de ce cambium génère le bois (xylème secondaire) et le liber (phloème secondaire).
Note :
Chez les gymnospermes (conifères), la croissance radiale est similaire à celle des dicotylédones. Un processus de croissance radiale différent existe aussi chez les fougères arborescentes.
Ramification
Ramification sympodiale dichasiale du lilas (Syringa vulgaris).
Ramification monopodiale de sapins rouges (Abies magnifica), les rameaux les plus jeunes et les plus courts sont proches du sommet (port pyramidal).
La tige peut rester entière, comme le chaume des graminées ou le stipe des palmiers, mais le plus souvent elle se développe en une structure plus ou moins ramifiée.
Les ramifications terminales (ou apicales) résultent de la division du méristème apical directement au niveau du dôme méristématique (ou de la cellule apicale). Selon que cette division entraîne la formation de deux axes ou plus, on parle respectivement de dichotomie ou de polytomie. Les embranchements peuvent être de taille égale (isotonie) ou non (anisotonie). C'est un mode de ramification archaïque qui se rencontre chez les bryophytes et les ptéridophytes, il est très rare chez les angiospermes et les gymnospermes.
Les ramifications latérales d'une tige sont dues au développement de ses bourgeons axillaires qui forment de nouveaux axes à une certaine distance du sommet. Ces rameaux latéraux peuvent former des rameaux de second ordre, qui à leur tour pourront en porter du troisième ordre, etc. C'est le mode de ramification le plus répandu chez les végétaux vasculaires. Il en existe deux variantes :
Dans les ramifications monopodiales, l'axe porteur grandit continuellement à partir de son bourgeons terminal (croissance indéfinie du méristème apical). Chaque axe est issu d'un unique méristème. On retrouve ce mode de ramification dans les inflorescences indéfinies de type grappe.
Dans les ramifications sympodiales, le bourgeon terminal de l'axe porteur ralentit ou arrête sa croissance (avortement du méristème apical ou différenciation en vrille, inflorescence ou fleur...). Un ou plusieurs bourgeons axillaires prennent le relai pour édifier autant de rameaux latéraux. Selon que l'on obtient un, deux ou plusieurs nouveaux axes, on parle respectivement de sympode monochasial, dichasial (gui, lilas), ou polychasial. Chaque segment d'un axe est issu d'un méristème différent. On retrouve ce mode de ramification dans les inflorescences définies de type cyme.
Ces deux modes de ramifications latérales ne sont pas exclusifs, certaines espèces peuvent être monopodiales jusqu'à la floraison, puis sympodiales (tomate).
Port des plantes
Arborescent
Buissonnant
Herbacée
Lianes
Fastigié
Pyramidal
Différents types de tiges
Tiges aériennes
cladodes
chaume des graminées
stipe (exemple : les palmiers)
tiges caulescentes
tiges succulentes
stolon
tige radicante
rameaux spécialisés épines vrilles
épines
vrilles
Tiges souterraines
Rhizomes
Les Rhizomes poussent horizontalement ou obliquement dans le sol. Ils portent des racines ou seulement un bouquet de feuilles.
Tubercules
Ces tiges se développent dans le sol, se remplissent de matières nutritives ils portent de bourgeons appelés "yeux"
Bulbes
Leur courte tige est appelée plateau. Ils portent a leur partie supérieur un bourgeon terminale entouré de feuilles réduites à des écailles charnues imbriquées, gorgées de matière nutritives.
Tiges Aquatiques
Coupe transversale de la tige d'une plante aquatique (Pontederia cordata). Noter les larges espaces intercellulaires remplis d'air pour assurer la flottabilité.
Les tiges des espèces qui vivent immergées dans l'eau (hydrophytes) ont une organisation particulière qui leur permet d'absorber directement l'eau, le dioxyde de carbone et l'oxygène, ainsi que des sels nutritifs. Ainsi, certaines plantes aquatiques telles que celles des genres Ceratophyllum, Utricularia et Wolffia, ne possèdent pas de racines qui leur seraient inutiles pour se nourrir. La paroi cellulaire des cellules épidermiques des tiges de ces plantes est recouverte par une mince cuticule perméable aux gaz, à l'eau et aux solutés. La présence de tissus de soutien n'est pas nécessaire en raison de la poussée d'Archimède. D'autre part, la plupart des plantes aquatiques présentent un développement remarquable des espaces intercellulaires, qui, en enfermant de l'air, améliorent la flottabilité ainsi que la diffusion de gaz dans la plante (voir aérenchyme).
Port de la tige
Dressée : la tige est suffisamment robuste pour se développer à la verticale.
Montante : concerne souvent des plantes dont la souche est vivace et robuste mais dont les tiges aériennes sont grêles et herbacées.
Couchée ou rampante : les tiges sont étalées au sol et ne montent pas ou peu. On parle également de plantes prostrées.
Volubile : entoure un support pour y prendre appui.
Grimpante : se fixe sur un support par des crampons qui sont des racines adventives ou par des vrilles, qui sont des feuilles transformées.
 词典释义:
词典释义:
 ,梗:
,梗: ,柄;【采】钻
,柄;【采】钻 :
:


 ;
;
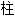 形的,
形的, 筒形的;
筒形的; , 柄 2. n. f 【植物学】梗 n. f. 【采】钻
, 柄 2. n. f 【植物学】梗 n. f. 【采】钻
 :~du piston活塞
:~du piston活塞
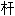 ~d'une colonne
~d'une colonne
 身
身
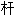 , 活塞
, 活塞 ; 调整
; 调整 ; 拉
; 拉 ; 茎; (苗)树干
; 茎; (苗)树干




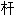






 、板、轨]
、板、轨]




 补心
补心 式顿钻钻井
式顿钻钻井
 接头
接头 阀[门]; 提升阀[门]; 钻
阀[门]; 提升阀[门]; 钻 浮阀[门]
浮阀[门]
 向
向 (压力壳端盖)
(压力壳端盖)