Dinosauria
Dinosauria Reconstitutions squelettiques de dinosaures non-aviens appartenant aux principaux clades de dinosaures. Dans le sens des aiguilles d'une montre et depuis le sommet gauche : Tyrannosaurus (Theropoda), Camarasaurus (Sauropodomorpha), Stegosaurus (Stegosauria), Triceratops (Marginocephalia), Maiasaura (Ornithopoda) et Gastonia (Ankylosauria). Classification Règne Animalia Embranchement Chordata Sous-embr. Vertebrata Super-classe Tetrapoda Classe Sauropsida Sous-classe Diapsida Division Archosauria Clade Ornithodira Super-ordre Dinosauria Owen, 1842 Clades de rang inférieur Saurischia † Herrerasauridae † Sauropodomorpha † Prosauropoda † Sauropoda Theropoda † Coelophysoidea † Ceratosauria Tetanurae † Ornithischia † Heterodontosauridae † Genasauria † Thyreophora † Cerapoda † Ornithopoda † Marginocephalia
Les dinosaures, du grec ancien δεινός / deinόs (« terriblement grand ») et σαῦρος / saûros (« lézard »), forment un clade extrêmement diversifié de vertébrés diapsides représentés actuellement par les oiseaux. Ce sont des archosaures ovipares, ayant en commun une posture érigée et partageant un certain nombre de synapomorphies telles que la présence d'une crête deltopectorale allongée au niveau de l'humérus et un acetabulum perforant le bassin. Présents dès la fin du Carnien (premier étage du Trias supérieur), il y a environ 230 Ma, ils disparaissent presque entièrement lors de la crise Crétacé-Tertiaire il y a 66 Ma. Seuls les oiseaux, qui émergent de petits dinosaures théropodes du Jurassique moyen, survivent à cette extinction, ce qui leur a permis de prospérer et se diversifier considérablement durant le Cénozoïque. Avec les oiseaux, les dinosaures forment aujourd'hui un des groupes de vertébrés les plus florissants, et les tétrapodes les plus diversifiés taxonomiquement et morphologiquement, leurs espèces étant actuellement plus nombreuses que celles des mammifères.
Si l'on ne prend pas en considération les oiseaux, les dinosaures dits « non-aviens » constituent un groupe de vertébrés majoritairement terrestres qui connut un succès évolutif considérable au Mésozoïque, dominant les faunes continentales entre le Trias supérieur et le Crétacé supérieur pendant plus de 160 Ma. Présents sur l'ensemble des continents dès la fin du Trias, ils comprennent des formes très diverses d'animaux terrestres et volants, bipèdes et quadrupèdes, carnivores et herbivores, ayant développé toute une série d'innovations squelettiques et tégumentaires telles que des cornes, des crêtes, des plaques et des plumes. Les dinosaures non-aviens comptent parmi eux les animaux les plus grands et les plus lourds ayant existé sur la terre ferme. Néanmoins, un grand nombre de dinosaures non-aviens ne dépassait pas la taille d'un être humain et certains d'entre eux étaient plus petits qu'une poule.
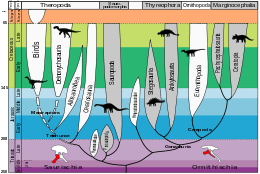
La classification des dinosaures distingue deux grands clades selon la morphologie de leur bassin : les Ornithischia et les Saurischia. Les Ornithischia (ou Ornithischiens) ne comprennent que des dinosaures herbivores que les paléontologues divisent en trois groupes majeurs, les Ornithopoda qui regroupent des dinosaures majoritairement bipèdes dont les fameux dinosaures à « bec de canard » (ou Hadrosauridae), les Marginocephalia qui incluent des dinosaures à collerette et à dôme osseux sur le haut de la tête (respectivement les Ceratopsia et les Pachycephalosauria), et enfin les Thyreophora qui englobent des dinosaures quadrupèdes surmontés d'armures, de piques et de plaques osseuses sur le dos et la queue (les Ankylosauria et les Stegosauria). Les Saurischia (ou Saurischiens) sont divisés en deux clades bien distincts, les Theropoda qui comprennent des dinosaures bipèdes et l'entièreté des dinosaures carnivores et piscivores ainsi que les dinosaures à plumes et les dinosaures volants (les oiseaux dans leur grande majorité), et les Sauropodomorpha, des animaux généralement quadrupèdes et de très grande taille munis d'un long cou, d'une petite tête et d'une longue queue.
Le terme Dinosauria fut proposé par le paléontologue anglais Richard Owen en avril 1842. Dès le milieu du XIX siècle et jusque dans les années 1970, les dinosaures furent considérés par les scientifiques comme des lézards géants, des reptiles à sang froid, patauds et lents, ayant disparu à la fin du Mésozoïque victimes de leur stupidité. Amorcée par le paléontologue américain John Ostrom, en 1969, la « renaissance des dinosaures » se caractérise par un regain d'intérêt pour l'étude des dinosaures, qui furent dès lors reconnus comme des animaux actifs, probablement endothermes même si toutes les études ne sont pas d'accord, et ayant des comportements sociaux complexes, bien distincts de nos reptiles actuels.
Les dinosaures suscitèrent dès leur découverte un grand intérêt auprès du public et les reconstitutions de squelettes dévoilées à l'occasion des expositions ont toujours été des attractions majeures dans les musées du monde entier. Les dinosaures sont d'ailleurs devenus partie intégrante de la culture populaire aux XXetXXI siècles, figurant dans une pléthore de livres et des films à succès tels que Jurassic Park et L'âge de glace 3. Aujourd'hui, si une « dinomania » touche notamment certains enfants, la curiosité populaire ne s'est jamais démentie et les nouvelles découvertes régulièrement rapportées par les médias entretiennent une certaine fascination pour ces animaux. Le mot « dinosaure » est largement passé dans le langage courant et son usage souvent inconsidéré tend à attribuer faussement ce terme à d'autres animaux disparus comme les ptérosaures, les reptiles marins (mosasaures, plésiosaures, etc.) et les reptiles mammaliens comme Dimetrodon qui n'étaient pas des dinosaures.
Étymologie
Le paléontologue anglais Sir Richard Owen (1804-1892), inventeur du terme Dinosauria.
Le substantif masculin dinosaure est emprunté, par l'intermédiaire de l'anglais dinosaur, au latin scientifique moderne dinosaurus.
Le taxon Dinosauria a été introduit et défini par le paléontologue britannique Richard Owen en 1842 afin de regrouper une « tribu ou sous-ordre distinct de Reptiles Sauriens ». Le terme dérive de deux racines grecques δεινός, deinos, signifiant, selon Owen, « effroyablement grand », et σαύρα, sauros, voulant dire « reptile » ou « lézard ». Dès l'apparition du nom, la première racine deinos fut incorrectement traduite par « terrible » ou « effrayant », bien qu'Owen n'utilisait pas cette racine comme un adjectif mais bien sa forme superlative, comme l'eût employé Homère dans l'Iliade, afin de révéler la nature impressionnante de ces animaux. Le paléontologue anglais incluait alors dans ce nouveau groupe de reptiles au moins trois genres, Megalosaurus, Iguanodon et Hylaeosaurus, tous trois caractérisés, entre autres, par un large bassin (sacrum) formé de cinq vertèbres ankylosées, la grande hauteur des épines neurales des vertèbres dorsales et la double articulation des côtes des vertèbres. Trois autres dinosaures préalablement nommés, le sauropode Cetiosaurus, le théropode Poekilopleuron et le sauropodomorphe basal Thecodontosaurus, ne furent cependant pas classés parmi les Dinosauria par Owen qui les considérait alors comme des reptiles indéterminés.
Définition
Définition phylogénétique
Squelette de Triceratops à l'American Museum of Natural History de New York.
Afin de distinguer les dinosaures de leurs plus proches cousins (les dinosauromorphes), les scientifiques modernes, qui définissent à présent des groupes d'organismes sur base de leurs ancêtres plutôt que sur la possession des caractères anatomiques particuliers, ont défini les dinosaures comme étant les « membres du clade le moins inclusif comprenant Triceratops horridus et Passer domesticus » (le moineau domestique).
Passer domesticus, le moineau domestique.
Cette définition, dite phylogénétique, a besoin d'un contexte phylogénétique et ne peut être comprise qu'en utilisant un cladogramme, ou arbre phylogénétique, un arbre schématique illustrant les liens de parentés entre les êtres vivants et les groupes au sein desquels ils se classent. Ainsi, un animal est un dinosaure s'il se positionne, dans un cladogramme, dans ce groupe d'organismes comprenant à la fois le Triceratops, le moineau, et l'ensemble des descendants de leur ancêtre commun. En d'autres termes, toute espèce descendant de cet ancêtre particulier est par définition un dinosaure. Cet ancêtre peut être positionné dans le cladogramme en trouvant le Triceratops et le moineau (Passer) puis en suivant leurs branches jusqu'à ce qu'elles se rejoignent en un nœud particulier correspondant au nœud du clade des dinosaures.
Des variantes de cette définition phylogénétique des dinosaures existent dans la littérature. Fernando Novas fut le premier à donner une définition du taxon Dinosauria, un clade incluant l'ancêtre commun des Herrerasauridae et des Saurischia + Ornithischia, nommant le clade comprenant les Saurischia et Ornithischia Eudinosauria, un taxon qui n'a jamais été repris par la suite. Un an plus tard, Kevin Padian et Cathleen May définirent le clade des Dinosauria comme étant l'ensemble des descendants du plus récent ancêtre commun des oiseaux et de Triceratops, la définition actuellement reconnue des dinosaures, reprise parfois de manière légèrement différente par d'autres auteurs comme le célèbre paléontologue américain Paul Sereno (Triceratops, Neornithes, leur plus récent ancêtre commun et l'ensemble de ses descendants), Olshevsky, qui souhaitait faire honneur à l'inventeur des dinosaures Richard Owen (le plus récent ancêtre commun de Megalosaurus et Iguanodon et l'ensemble de ses descendants) et Fraser et ses collègues (Triceratops (représentant les Ornithischia) plus Aves (représentant les Saurischia) et l'ensemble des descendants de leur dernier ancêtre commun).
Illustration schématique représentant la définition phylogénétique du clade des dinosaures.
L'ensemble de ces définitions permet de regrouper des animaux définis comme étant des dinosaures (Dinosauria = Saurischia + Ornithischia) et comprenant les théropodes (dinosaures bipèdes, parfois carnivores, incluant les oiseaux), les sauropodomorphes (dinosaures herbivores majoritairement quadrupèdes munis d'un long cou), les ornithopodes (dinosaures herbivores majoritairement bipèdes comprenant les dinosaures à « bec de canard »), les ankylosauriens (dinosaures herbivores quadrupèdes munis d'armures osseuses sur le dos), les stégosauriens (dinosaures herbivores quadrupèdes portant des plaques osseuses et parfois des épines sur le dos), les cératopsiens (dinosaures herbivores majoritairement quadrupèdes possédant un crête osseuse à l'arrière du crâne et parfois des cornes sur le haut de la tête) et les pachycéphalosauridés (dinosaures herbivores bipèdes au crâne épaissi).
Il y a un consensus presque total chez les paléontologues sur le fait que les oiseaux sont les descendants des dinosaures, et plus précisément des dinosaures théropodes appartenant au clade des Maniraptora. Suivant la définition phylogénétique, les oiseaux sont donc des dinosaures et les dinosaures n'ont par conséquent pas disparu. Néanmoins, dans le langage courant, le mot « dinosaure » n'inclut pas les oiseaux. Par souci de clarté, cet article utilise ainsi le mot « dinosaure » comme synonyme de « dinosaure non-avien ».
Caractères anatomiques identifiant les dinosaures
Les premiers scientifiques qui étudièrent les dinosaures ne leur avaient pas donné une telle définition puisque la théorie de l'évolution, impliquant la notion d'ancêtre commun, était vue à leur époque comme une hérésie. Ainsi, suivant la coutume de son époque, Richard Owen définit les dinosaures comme étant des animaux possédant un certain nombre de caractères anatomiques particuliers. Ces derniers sont actuellement nommés synapomorphies, des caractères dérivés, ou nouveautés évolutives, hérités d'un ancêtre commun et unissant l'ensemble des membres d'un clade. Les dinosaures possèdent plusieurs synapomorphies listées ci-dessous. Ces caractères dérivés partagés par l'ensemble des dinosaures permettent, non pas de définir les dinosaures, mais de les identifier, comme un médecin utiliserait une liste de symptômes pour diagnostiquer une maladie particulière.
Crâne d'Herrerasaurus (Saurischia) en vue dorsale montrant l’extension antérieure de la zone d'attache des muscles temporaux au niveau de la partie antérieure de la fosse temporale supérieure.
Musculature temporale s'étendant antérieurement sur le toit du crâne (= fosse supratemporale présente dans la partie antérieure de la fenêtre supratemporale). Les muscles adducteurs mandibulaires, des muscles temporaux permettant la mastication chez les vertébrés, sont particulièrement larges et développés chez les dinosaures. Ainsi, la zone osseuse où s'attachent ces muscles sur le haut du crâne, dans la partie antérieure de la fosse temporale, est particulièrement étendue, alors qu'elle est courte chez les autres archosaures et les cousins très proches des dinosaures comme Silesaurus.
Processus postérieur du jugal bifurque pour s'articuler avec le quadratojugal. Chez les archosaures autres que les dinosaures, le processus du jugal s'étendant postérieurement et s'articulant avec le quadratojugal est unique alors que chez les dinosaures, il est divisé en deux processus distincts.
Présence d'épipophyses chez les vertèbres cervicales antérieures (et non axiales). Les épipophyses sont des projections osseuses positionnées dans la partie postérodorsale des épines neurales des vertèbres cervicales (sur la partie dorsale des postzygapophyses plus précisément). Elles permettent à certains muscles du cou de s'attacher afin de renforcer le cou et le dos. Chez les dinosaures, ces muscles devaient être particulièrement développés et capables d'une large variété de mouvements.
Humérus de Stegosaurus (Stegosauridae) présentant la crête deltopectorale allongée et développée.
Une crête deltopectorale allongée au niveau de l'humérus (occupant plus de 30 % la longueur de l'humérus). La crête deltopectorale est une ride osseuse située sur la partie antérieure de l'humérus. Elle permet aux muscles deltoïdes de l'épaule et pectoralis de la poitrine de s'insérer. Chez les archosaures autres que les dinosaures, cette crête est discrète, alors qu'elle est longue et proéminente chez les dinosaures, témoignant de la grande mobilité et de la puissance des membres antérieurs chez les dinosaures.
Bassin d'Allosaurus (Theropoda) montrant l'acétabulum ouvert. Un acetabulum ouvert au niveau du bassin (= marge ventrale de l'acétabulum de l'ilion concave). L'acetabulum est une cavité osseuse du bassin où s'insère et s'articule la tête du fémur. Chez l'homme, cette surface articulaire est circulaire afin de recevoir la tête arrondie du fémur, permettant une grande amplitude de mouvement. Chez les dinosaures, et contrairement à leurs proches cousins archosaures, l'acetabulum est ouvert, ou perforé. Une perforation de l'acetabulum témoigne d'une posture dressée, où les membres postérieurs se situent sous le corps. Chez les animaux à posture semi-dressée ou rampante, la surface médiale de l'acetabulum est renforcée par de l'os afin de dissiper les contraintes soumises par le poids de l'animal qui se propagent médialement vers le bassin, depuis la tête du fémur. Chez les dinosaures qui ont une posture dressée, la contrainte soumise par le poids de l'animal et exercée sur la surface articulaire de l'acetabulum est, non plus médiale, mais dorsale. Ainsi, la surface médiale osseuse de l'acetabulum, devenue inutile chez les archosaures à posture dressée, s'est vue disparaître au cours de l'évolution.
Le quatrième trochanter du fémur forme une crête forte et asymétrique. Le quatrième trochanter est une crête osseuse située sur la partie postérieure du fémur et présent chez l'entièreté des archosaures. Ce trochanter permet l'insertion des muscles caudo-fémoraux de la jambe pour la locomotion. Chez la plupart des archosaures, le quatrième trochanter est peu développé et symétrique alors que chez l'entièreté des dinosaures, ce trochanter est protubérant et asymétrique, avec une partie distale plus étendue que la partie proximale. Bien que l'utilité d'un quatrième trochanter asymétrique est incertaine, le développement important de cette arête osseuse témoigne de membres postérieurs puissants et doués de mouvements efficients.
Fémurs d'Olorotitan (Hadrosauridae) illustrant la position du quatrième trochanter, fortement développé et asymétrique chez les dinosaures.
Une facette articulaire de la fibula occupant moins de 30 % la largeur de l'astragale. L'astragale et le calcanéus sont les os proximaux du tarses (des os du pied) chez les archosaures. Chez les dinosaures et ses proches cousins, l'astragale est beaucoup plus grand que le calcanéum et les deux os sont fermement articulés entre eux, ainsi qu'avec le tibia et la fibula, l'ensemble de ces quatre os formant ainsi un complexe fonctionnel unique ne permettant aucun mouvement de rotation entre ceux-ci. Chez ces animaux, les mouvements entre la jambe et la patte se font entre le fémur, le complexe tarsal proximal (tibia + fibula + astragale + calcanéum) et le pied lui-même, une condition dite « mésotarsale » partagée par la lignée avienne des archosaures (Avemetatarsalia), qui se distingue de celle des pseudosuchiens où l'astragale et le calcanéum sont de taille semblable et où les deux os s'articulent de manière mobile. Les dinosaures se distinguent des autres métatarsaliens par un contact très réduit entre la fibula et l'astragale puisque la surface de contact entre les deux os n'occupe seulement que 30 % de la largeur de l'astragale.
Et selon Nesbitt (2011) :
Les exoccipitaux ne se rejoignent pas le long de la ligne du milieu de la surface de la cavité endocrânienne.
Les radius occupent moins de 80 % la longueur la longueur de l'humérus.
La crête cnémiale du tibia s'arque antérolatéralement.
Présence d'une ride longitudinale distincte sur la face postérieure de la terminaison distale du tibia.
La surface d'articulation avec la fibula est concave sur le calcanéus.
Absence d'un os postfrontal au niveau du crâne.
Plaques sternales ossifiées et doubles.
Doigts IV et V de la main réduits.
Trois phalanges ou moins au niveau du quatrième doigt de la main.
Présence d'un foramen post-temporal.
Trois dents au niveau du prémaxillaire.
Fosse nasale du prémaxillaire s'étendant dans le coin antéroventral de la narine.
L'échancrure ventrale de la tête du fémur est concave.
Absence d'un sillon postérieur sur l'astragale.
Absence d'une tubérosité sur le calcanéum.
Absence d'interclavicules.
Une crête/fosse brevis sur la surface ventrale de l'ilion. La plupart des dinosaures possèdent une fosse distincte sur la surface latérale ou ventrale de l'ilion. C'est à cet endroit que s'attachaient une partie de la musculature caudofémorale dites "caudofemoralis brevis". La distribution de cette fosse est complexe puisqu'il n'est pas certain qu'il s'agisse d'un caractère homologue. Ainsi, les dinosaures basaux Herrerasaurus et Plateosaurus ne possèdent pas une telle fosse, au contraire de leur proche cousin Silesaurus et des dinosaures basaux Eocursor, Lesothosaurus, Panphagia et Saturnalia.
Trois vertèbres sacrées ou plus. La plupart des archosaures tels que les dinosauromorphes Lagerpeton et Marasuchus possèdent deux vertèbres sacrées au niveau du bassin. La grande majorité des dinosaures comprennent trois ou plus de trois vertèbres sacrées mais les saurischiens basaux Herrerasaurus et Staurikosaurus montrent deux vertèbres sacrées alors que Silesaurus, un taxon sœur des dinosaures en possèdent trois également.
Position dorsale de l'ectoptérygoïde par rapport à la crête transversale du ptérygoïde.
Cavité glénoïde orienté postéro-ventralement au niveau de la scapula (omoplate) et du coracoïde.
Contact entre le pubis et l'ischion réduit.
Lamine médioventrale de l'ischion réduite.
Tête du fémur tournée vers l'intérieur.
Partie proximale du fémur avec une tubérosité médiale réduite.
Présence d'un trochanter antérieur au niveau du fémur.
Processus descendant du tibia s'insérant postérieurement par rapport au processus ascendant de l'astragale.
Calcanéum proximal plat et concave.
Présence d'une cheville métotarsale.
Métatarses II et IV de longueur subégale.
Terminaison distale du métatarse IV et plus long que large.
Description
Généralités
Rhamphorhynchus n'est pas un dinosaure mais un ptérosaure, un archosaure volant (vue d'artiste).
Bien que nécessaire, la liste des traits anatomiques énoncée ci-dessus et permettant de différencier un dinosaure d'un autre animal est rébarbative et emploie un grand nombre de termes anatomiques complexes et peu intéressants pour un public non-spécialisé. Ainsi, un animal d'origine archosaurienne ayant vécu au Mésozoïque, ne volant pas et se différenciant des reptiles par une posture « érigée en contrefort » est très souvent, un dinosaure (non-avien).
Le Dimetrodon n'est pas un dinosaure mais un thérapside ayant vécu au Permien, bien avant les premiers dinosaures (vue d'artiste).
Puisque les dinosaures sont des archosaures, les mammifères disparus tels le Mammouth, ne sont pas des dinosaures, de même que tout animal ayant vécu avant le Trias comme le Dimetrodon, un thérapside du Permien. Les dinosaures furent des animaux majoritairement terrestres et seuls quelques dinosaures à plumes proches de la lignée avienne comme Microraptor étaient capables de voler et d'occuper ainsi la niche écologique des airs. Par conséquent, les animaux aquatiques ayant vécu à la même époque que les dinosaures, comme les plésiosaures, les ichtyosaures et les mosasaures, n'étaient pas des dinosaures. De la même manière, et bien qu'ayant également vécu uniquement au Mésozoïque également, les ptérosaures, des proches cousins des dinosaures, exclusivement volants et dont l'aile est formée par une voile de peau soutenue par le quatrième doigt de la main qui s'est considérablement allongé, ne sont pas des dinosaures. Puisque les dinosaures possédaient des membres dressés, maintenus sous le corps, les animaux proches des dinosaures à posture semi-dressée ou rampante comme les reptiles, tels les crocodiles ou les lézards, ne sont pas non plus des dinosaures.
Batrachotomus, un rauisuchien à posture « érigée en pilier » (vue d'artiste).
La posture dressée n'est pas une exclusivité dinosaurienne au sein de la lignée archosaurienne puisqu'elle est également partagée, en plus des proches cousins des dinosaures les dinosauromophes, par d'autres archosaures contemporains des premiers dinosaures : les Rauisuchia. La posture érigée de ces archosaures est néanmoins différente de celle des dinosaures (et des mammifères) dans la configuration anatomique du bassin. Chez les dinosaures (et les ornithodires d'une manière générale), la stature dressée fut acquise en gardant une position verticale des os du bassin et en tournant la tête du fémur médialement afin qu'elle s'articule dans la cavité de l'acetabulum qui s'ouvre latéralement, une posture dite « érigée en contrefort » ('buttress-erect' posture). Chez les rauisuchiens, la stature érigée, qui est apparue indépendamment de celle des dinosaures, s'est développée grâce à une rotation de l'ilion vers une position horizontale, orientant ainsi l'acetabulum presque ventralement. Cette configuration est connue sous le nom de posture « érigée en pilier » ('pillar-erect' posture).
Posture des archosaures : semi-érigée chez les crocodiliens, érigée en contrefort chez les dinosaures, et érigée en pilier chez les rauisuchiens.
Il est nettement plus difficile de différencier les tout premiers dinosaures de leur proches cousins les dinosauromorphes puisque ces derniers sont également des archosaures bipèdes à posture « érigée en contrefort » ayant vécu à la même époque que les premiers dinosaures. Comme le montre la liste de caractères anatomiques propre aux dinosaures donnée ci-dessus, les dinosaures ne se différencient véritablement de leurs proches cousins que par une posture dressée plus rapide et plus effective, où les muscles du cou et des membres antérieurs et postérieurs furent plus développés et plus puissants, permettant aux premiers dinosaures d'avoir une bipédie effective et un mode de vie très actif.
Silesaurus, un dinosauromorphe, est un des cousins les plus proches des dinosaures. Il ne se différencie de ces derniers que par une posture dressée moins effective et moins rapide (vue d'artiste).
Seule une posture dressée et des membres antérieurs et postérieurs pourvus de muscles puissants permettent de caractériser l'ensemble des dinosaures, qui furent de taille et de morphologie extrêmement diverses. En effet, les dinosaures, comprennent des animaux bipèdes et quadrupèdes, carnivores et herbivores, avec ou sans dents, à écaille, à poils (des proto-plumes) ou à plumes, aux membres antérieurs très développés ou très courts, des cous démesurément longs ou presque absents, et munis ou non de becs, collerettes, armures osseuses, plaques, cornes, épines, voiles, crêtes et dômes osseux.
Crâne et mâchoire
Tête d'Acrocanthosaurus, un Carcharodontosauridae.
Le crâne et la mâchoire des dinosaures sont certainement les parties du squelette dont la forme est la plus variée. En effet, la morphologie de la tête, que ces deux entités anatomiques constituent, est directement liée au régime alimentaire, si bien que les dinosaures qui se nourrissaient de plantes coriaces, de feuilles, de viandes, de poissons ou filtraient les eaux pour se sustenter avaient des crânes et des mâchoires de forme très différente. Ainsi, les dinosaures théropodes qui comprennent des prédateurs puissants comme Tyrannosaurus, Acrocanthosaurus et Torvosaurus avaient des crânes massifs et robustes munis de dents tranchantes dentelées, alors que leur proches cousins piscivores les Spinosauridae comme Irritator et Spinosaurus avaient un crâne étroit et allongé portant de dents coniques dépourvues de denticules. Beaucoup de théropodes étaient également exclusivement herbivores et possédaient des têtes à bec édenté comme certains Oviraptoridae tel que Citipati ou des mâchoires à bec munies de petites dents triangulaires comme les Therizinosauroidea tel Erlikosaurus.
Tête de Stegoceras, un Pachycephalosauridae.
Tête de Corythosaurus, un Hadrosauridae.
Parmi les dinosaures ornithischiens, qui furent exclusivement herbivores, le crâne était également souvent muni de becs osseux comme la plupart des cératopsiens et des hadrosaures dont les mâchoires portaient de véritables batteries dentaires. Néanmoins, la forme du crâne des ornithischiens, en plus d'être liée au régime alimentaire, variait également fortement suivant les attributs de protections ou de parades comme les cornes, les dômes osseux, les crêtes et les collerettes qu'il portait. Ainsi, la tête des cératopsiens comme Styracosaurus, Pachyrhinosaurus et Triceratops, qui fut munie d'une collerette (courte ou longue) et parfois de cornes nasales, frontales ou encore de cornes à l'extrémité de la collerette, se distinguait fortement de la tête épaisse et étroite des pachycéphalosauridés comme Stegoceras ou Pachycephalosaurus dont le crâne était épaissi et parfois bombé à son sommet. De même, la tête des hadrosaures, souvent pourvue de longues crêtes osseuses à l'extrémité du crâne comme chez Corythosaurus et Parasaurolophus, ne pouvait être confondue avec la tête épaisse et courte des ankylosauridés comme Ankylosaurus et Euoplocephalus dont le sommet était protégé de protubérances osseuses et parfois rehaussé de courtes épines.
Crâne d'Euoplocephalus, un Ankylosauridae.
Malgré cette large variabilité de forme que déploie la tête des dinosaures, le crâne et la mâchoire sont toujours constitués des mêmes os et montrent toujours les mêmes ouvertures. En effet, les dinosaures sont des reptiles diapsides, des animaux dont le crâne est perforé par deux ouvertures crâniennes, ou fenêtres (également appelées fosses), dans la partie postérieure du crâne, la fenêtre temporale inférieure et la fenêtre temporale supérieure. De même, comme la majorité des vertébrés, les dinosaures possèdent une ouverture à l'emplacement de l’œil nommée orbite au milieu duquel se trouve l'anneau sclérotique permettant le support de l’œil, ainsi qu'une ouverture au niveau des narines, la fosse nasale d'où se terminaient les conduits respiratoires jusqu'aux narines. L'entièreté des saurischiens, et certains ornithischiens, montrent enfin une dernière fenêtre, généralement large, au niveau du crâne et située entre l'orbite et la fosse nasale. Cette ouverture, appelée fenêtre antéorbitaire, ce qui signifie "en avant de l'orbite", permettait le passage de plusieurs muscles reliant le toit du crâne aux mâchoires supérieures et inférieures. La mandibule est également pourvue d'une ouverture nommée fenêtre mandibulaire externe, présente chez la plupart des dinosaures.
Anatomie du crâne et de la mâchoire de Spinosaurus, un Spinosauridae.
Tête de Triceratops, un Ceratopsidae à trois cornes.
Les os formant le crâne et la mâchoire, en grande majorité double (présents du côté gauche et droit) puisque la tête est symétrique, sont nombreux et leur morphologie est également très diverse au sein des différents groupes de dinosaures. Seuls les plus caractéristiques sont ici exposés. Le prémaxillaire se situe à l'extrémité antérieure du crâne et porte des dents ou un bec corné appelé ramphothèque, ou les deux. Néanmoins, chez les hadrosaures, il existe deux prémaxillaires qui contribuent à la crête osseuse portée par bon nombre d'entre eux. Le maxillaire est également, chez les dinosaures dentés, un os porteur de dents qui constitue la majeure partie de la mâchoire supérieure. Les os formant le toit du crâne sont l'os nasal (délimitant en partie la fosse nasale et formant parfois des crêtes osseuses de toutes sortes), l'os frontal, et l'os pariétal et squamosal qui se trouvent à l'arrière de la tête. Ces deux derniers sont extrêmement développés chez la plupart des cératopsiens puisqu'ils forment la large crête permettant de protéger le cou de ces animaux. La mandibule est majoritairement constituée de l'os dentaire qui est le seul à porter des dents au niveau de la mâchoire inférieure. Chez les dinosaures à bec comme les cératopsiens, les stégosauridés et les hadrosauridés, l'extrémité antérieure de la mâchoire se termine par un os supplémentaire directement relié au dentaire, l'os prédentaire, qui est également renforcé par une ramphothèque chez ces animaux à bec. De la même manière, chez les cératopsiens le prémaxillaire de la mâchoire supérieure est prolongé par un os supplémentaire portant le bec corné et appelé os rostral.
Squelette post-crânien
Plaques dorsales losangiques d'Hesperosaurus, un Stegosauridae.
Si le squelette post-crânien (le squelette mis à part la tête) des dinosaures est moins différencié que le crâne et la mandibule, il n'en reste pas moins que sa morphologie est très variable au sein des différents clades de dinosaures. Directement lié à la locomotion et à l'alimentation, le squelette des dinosaures pouvait également déployer toute une série d'innovations squelettiques jouant des rôles de protection, de défense et de parade. Les dinosaures carnivores étaient ainsi exclusivement bipèdes afin de se déplacer avec agilité et rapidité pour attraper des proies. Leurs membres postérieurs furent nettement plus développés et robustes que les membres antérieurs, et les doigts de ces animaux se terminaient par des griffes acérées et parfois rétractiles. Au contraire, une majorité de dinosaures herbivores fut quadrupède et beaucoup d'entre eux se munirent de protubérances osseuses pour se protéger. Si la tête de certains Ceratopsidae déployaient des cornes, le corps des Stegosauridae portait des plaques osseuses triangulaires ou losangiques sur le haut du dos et parfois de longues épines à l'extrémité de la queue et sur les bas côté du ventre.
Bouclier osseux de Sauropelta, un Ankylosauridae.
Les Ankylosauridae adoptèrent une autre stratégie pour se prémunir des prédateurs. Chez ces animaux quadrupèdes, le corps était recouvert d'un bouclier osseux fait d'une imbrications de nodules, de piques et parfois de larges épines pointant vers les côtés. La queue de ces herbivores se terminait parfois également par une massue osseuse que l'animal était capable de balancer pour repousser les assaillants les plus téméraires. Un autre moyen de se protéger des prédateurs et vers lequel les Sauropodomorpha évoluèrent fut l'accroissement démesuré du corps, où le cou et la queue prirent des longueurs gigantesques. La grande dimension du cou permit à ces dinosaures quadrupèdes d'atteindre la cime des arbres les plus hauts pour se nourrir alors qu'une queue excessivement longue aida à contrebalancer le reste du corps et parfois également à jouer un rôle de fouet supersonique capable de claquer face aux carnassiers qui les attaquaient. Des nodules et épines osseuses se trouvaient également sur le dos de certains sauropodes titanosaures tels qu'Agustinia et Ampelosaurus, de même que certains théropodes carnivores comme Ceratosaurus. La longue queue de certains sauropodes se terminait également par une massue, comme le montre Shunosaurus et Omeisaurus, et parfois même de courtes épines telle la queue de Spinophorosaurus.
Squelette de Patagonykus (Alvarezsauridae) dont les membres antérieurs ne portent qu'un seul doigt.
Les dinosaures herbivores bipèdes, comme la plupart des Ornithopoda, des Pachycephalosauridae et un grand nombre de théropodes maniraptoriformes, comptèrent surtout sur la robustesse et la vélocité de leurs membres postérieurs pour échapper aux prédateurs, mais certains herbivores comme Iguanodon portèrent également de larges éperons au niveau des pouces afin de poinçonner le cou des prédateurs. Bien que la plupart des théropodes comprenaient trois à quatre doigts à chaque main, quelques-uns d'entre eux comme Tyrannosaurus n'étaient pourvus que de deux doigts développés et un troisième doigt vestigial totalement réduit. Par ailleurs, les membres antérieurs de certains théropodes Alvarezsauridae ne furent terminés que par un doigt unique muni d'une large griffe. Les membres antérieurs des théropodes furent généralement bien développés, parfois même robustes comme chez les Spinosauridae, mais certains d'entre eux au sein de la lignée des Abelisauridae, Tyrannosauridae et Alvarezsauridae montraient des membres antérieurs extrêmement courts, leur réduction au cours du temps témoignant parfois d'une absence d'utilité. Dinosaures herbivores et carnivores pouvaient également déployer une voile osseuse sous-tendue par des épines neurales hypertrophiées sur le haut du dos. C'est le cas des dinosaures ornithopodes Ouranosaurus et Hypacrosaurus ainsi que les dinosaures théropodes Acrocanthosaurus, Concavenator et Spinosaurus.
Gastralias (os du ventre) de Tyrannosaurus (Tyrannosauridae).
Une fois encore, cette large variabilité de forme du corps des dinosaures ne dérive que de la transformation d'un certain nombre d'os, et la toute grande majorité des os du squelette des dinosaures fut partagée par l'ensemble d'entre eux et configurée de la même manière. Ainsi, la colonne vertébrale comprend toujours un nombre (parfois variable) de vertèbres cervicales (vertèbres du cou), dorsales (du dos), sacrales (du bassin) et caudales (de la queue). La cage thoracique est protégée par les côtes (cervicales et dorsales) et des os fins et parallèles nommés gastralias au niveau du bas-ventre, et les petits os situés sous les vertèbres caudales et permettant aux muscles de la queue de s'y attacher sont appelés chevrons ou arcs hémaux. Les membres antérieurs incluent un humérus (partie proximale de l'avant-bras), un radius et une ulna (partie distale de l'avant-bras) et enfin les os de la main que sont les carpes, métacarpes et phalanges. De même, les membres postérieurs se divisent en fémur (partie proximale de la jambe), tibia et fibula (partie distale de la jambe) et enfin des os du pied, à savoir les tarses, métatarses et phalanges. Le complexe scapulaire formant l'épaule comprend la scapula et le coracoïde ainsi que deux petits os, le sternum et la clavicule (cette dernière étant fusionnée chez les théropodes pour former un os en forme de 'V' nommé furcula) et le bassin se constitue d'un ilion, d'un ischion et d'un pubis. Les piques, plaques et nodules que portent certains dinosaures sont enfin des ostéodermes, des protubérances osseuses dérivant de l'ossification de certaines parties du derme.
Organes internes
Le spécimen Scipionyx samniticus montrant les restes minéralisés des organes internes exceptionnellement préservés.
Le cœur fossilisé du Thescelosaurus «Willo» qui semble se révéler n'être qu'une concrétion rocheuse.
Si l'anatomie osseuse des dinosaures est relativement bien connue grâce à la découverte de leurs os fossilisés, il en est tout à fait autrement de leur anatomie interne. En effet, les organes internes que sont par exemple le cerveau, les organes respiratoires et digestifs (poumons, estomacs, intestins, etc.) et les muscles, se dégradent rapidement une fois l'animal mort et seuls des cas exceptionnels de dépôt rapide et de fossilisations permettent à ces parties molles du corps d'être préservés dans la roche. Un des plus bels exemples est donné par le théropode juvénile Scipionyx dont les organes et tissus mous ont été particulièrement bien conservés. La préservation unique des ligaments, des cartilages et des muscles du corps, des vaisseaux sanguins et de l'entièreté des organes digestifs chez ce petit coelurosaure a permis d'apporter des éléments cruciaux quant aux connaissances sur l'anatomie générale des dinosaures qui semble se rapprocher beaucoup plus de celle des oiseaux que des reptiles contemporains. Bien qu'à un degré inférieur de préservation, des restes de viscères et de muscles ont été également fossilisés chez d'autres dinosaures tels que Mirischia préservant des traces d'intestins, Santanaraptor et Pelecanimimus montrant des restes de peau et de muscles et enfin plusieurs exemples d'hadrosauridés superbement momifiés comme Brachylophosaurus (nommé «Leonardo») et Edmontosaurus (nommé «Dakota») ayant tous deux préservés des restes tégumentaires, ligamentaires et viscéraux. La découverte d'un cœur fossilisé, qui semble avoir été divisé en quatre chambres (deux aortes et deux ventricules) comme celui des mammifères et des oiseaux, a été également reporté chez l'ornithopode Thescelosaurus. Un examen minutieux de ce cœur pétrifié, qui aurait été conservé par saponification et transformation en goethite selon certains, a révélé qu'il s'agit-là plus vraisemblablement d'une concrétion rocheuse.
Musculature du crâne et de la mandibule chez Spinosaurus.
La musculature des dinosaures a été particulièrement étudiée ces dernières années et les reconstitutions musculaires se basent moins sur les tissus musculaires préservés, qui restent très rares dans le registre fossile, mais sur les attaches musculaires visibles sur les os et une comparaison minutieuse avec l'anatomie musculaire des proches parents des dinosaures vivants de nos jours, à savoir les oiseaux et les crocodiles. Ainsi, les muscles du crâne et de la mâchoire, qui comprennent les muscles orbito-temporaux , palataux , temporaux et mandibulaires , ont été récemment reconstitués chez Edmontosaurus, Diplodocus et Majungasaurus. Quant aux muscles du cou, une reconstitution de leur anatomie a pu être donnée chez certains théropodes, marginocéphales et sauropodes. De la même manière, des données nouvelles sur la musculature des membres antérieurs et postérieurs chez les dinosaures ont été récemment apportées, par exemple chez Tyrannosaurus et les Dromaeosauridae, contribuant à développer de manière importante les connaissances sur la locomotion des dinosaures.
Anatomie crânienne et reconstitution de l'encéphale de Nigersaurus, et évolution du cerveau chez les sauropodomorphes.
Du fait de sa complexité, la boîte crânienne (ou neurocrâne) renfermant le cerveau a été pendant longtemps une des régions du squelette des dinosaures les moins connues. Néanmoins, les connaissances relativement faibles de son anatomie aux siècles derniers se sont considérablement améliorées au XXI siècle avec l'avènement du CT-scan. Cet outil utilisé dans les hôpitaux a permis de scanner avec précision et dans les moindres détails la cavité céphalique des boîtes crâniennes relativement bien conservées de nombreux dinosaures. Des reconstructions en 3 dimensions des hémisphères cérébraux, des bulbes olfactifs et des organes de l'oreille interne ont pu être ainsi faites chez plusieurs dinosaures comme Majungasaurus, Tyrannosaurus, Alioramus, Hypacrosaurus, Pachyrhinosaurus, Diplodocus, Camarasaurus et Nigersaurus. Ces études ont permis de comprendre beaucoup mieux l'anatomie du cerveau des dinosaures, mais également d'avoir une idée de leur intelligence et de leurs facultés sensorielles (ouïe, odorat, vue). L'on sait à présent que la morphologie du cerveau est très variable au sein du groupe des dinosaures et les ornithischiens, les sauropodomorphes et les théropodes basaux possédèrent généralement un cerveau proche des reptiles actuels dans sa configuration anatomique, alors que les théropodes dérivés avaient un cerveau ressemblant beaucoup plus à celui des oiseaux. Le cerveau des dinosaures peut se diviser en deux parties, le bulbe olfactif antérieurement et les hémisphères cérébraux au centre, auxquels s'ajoutent, de part et d'autre du cerveau, les 12 nerfs majeurs du cerveau (notés de I à XII), les veines cérébrales et enfin l'oreille interne, constitué du labyrinthe osseux. L'évolution du cerveau chez les théropodes correspond ainsi a un accroissement important du volume cérébral chez les coelurosaures, de même qu'un alignement du bulbe olfactif, des hémisphères cérébraux (télencéphale) et du rhombencéphale sur un même plan.
Peaux, poils et plumes
Empreinte de peau écailleuse du Ceratopsidae Centrosaurus
Les dinosaures furent classés pendant longtemps comme des reptiles couverts d'écailles tels nos crocodiles et lézards actuels. Cette vision a fortement changé de nos jours depuis la découverte de structures osseuses typiques des animaux homéothermes (à sang chaud) chez de nombreux dinosaures, et depuis que la province du Liaoning, en Chine, a livré dans les années 1990 et 2000 un grand nombre de fossiles de dinosaures couverts de poils et de plumes. Si certains dinosaures portaient ce genre de téguments filamenteux, d'autres semblent avoir eu la peau lisse, et d'autres encore avoir été couverts d'écailles ; plusieurs types de phanères on pu coexister, comme chez les oiseaux actuels. En effet, des traces d'écailles ont été découvertes chez de nombreux groupes de dinosaures comme les hadrosauridés (par exemple, Saurolophus, Brachylophosaurus, Edmontosaurus), les cératopsidés (p. ex. Centrosaurus, Chasmosaurus), les stégosauridés (p. ex., Gigantspinosaurus, Hesperosaurus), les ankylosauridés (p. ex. Scelidosaurus, Tarchia), les sauropodes (p. ex., Saltasaurus, Tehuelchesaurus) et les théropodes (p. ex., Carnotaurus, Juravenator). Il s'agit dans la plupart des cas d'impressions de peau dans le sédiment mais il existe des cas exceptionnels de peaux fossilisées dont la structure moléculaire a été préservée, comme les momies de dinosaures dont la plupart sont des hadrosaures.
Le duvet pileux du Compsognathidae Sinosauropteryx
Le dinosaure à plumes Caudipteryx
La très grande majorité des paléontologues s'accorde à présent à dire que les dinosaures, et plus exactement les théropodes, furent les ancêtres directs des oiseaux, et les preuves de plumes chez certains théropodes non-aviens ont grandement conforté cette théorie. En effet, l'évolution de la plume se suit très bien tout au long du clade des dinosaures, depuis les dinosaures relativement basaux portant un duvet pileux, jusqu'aux théropodes dérivés déployant de véritables rémiges. La présence de poils est avérée chez aux moins deux ornithischiens, l'Heterodontosauridae Tianyulong et le Psittacosauridae Psittacosaurus, et un certain nombre de théropodes dont le Megalosauridae Sciurumimus, le Tyrannosauroidea Dilong, les Compsognathidae Sinosauropteryx et Juravenator, et l'Alvarezsauridae Shuvuuia. Ces poils de structures très simples se différencient déjà des téguments filamenteux plus larges et plus longs du Therizinosauroidea Beipiaosaurus. L'Oviraptoridae Caudipteryx et le Dromaeosauridae Sinornithosaurus portent quant à eux des proto-plumes, des structures tégumentaires branchues comprenant une tige centrale à laquelle se joignent des filaments latéraux. Finalement, plusieurs coelurosaures dérivés dont les Oviraptoridae Caudipteryx et Similicaudipteryx, les Dromaeosauridae Microraptor et les Troodontidae Anchiornis furent couverts de plumes de type pennes tout à fait homologues à celles de nos oiseaux actuels. Bien qu'aucun fossile de Velociraptor n'ait conservé des empreintes de plumes, les os des membres antérieurs, et plus spécifiquement l'ulna, montrent de petites protubérances alignées de manière régulière le long de l'os et interprétées comme étant le lieu de fixation de plumes modernes (formée de rachis, vannes et barbules). Des structures similaires ont également été découvertes le long de l'ulna du Carcharodontosauridae Concavenator, un théropode beaucoup plus primitif que les maniraptoriformes à plumes, démontrant, s'il ne s'agit pas là de lieux d'attachement des muscles, la présence de rémiges le long des bras déjà chez ces théropodes relativement primitifs.
Les plumes du dinosaure Dromaeosauridae Microraptor
Bien que la plupart des dinosaures semble avoir porté uniquement des écailles, il n'est pas impossible que ceux-ci aient perdu la pilosité de leur ancêtres. En effet, les dinosaures à poils les plus primitifs appartiennent au groupe des Heterodontosauridae, des ornithischiens primitifs de petite taille, néanmoins la présence de poils est également avérée chez certains ptérosaures comme Sordes, des reptiles volants proches parents des dinosaures. Il n'est pas encore certain que les structures filamenteuses des ptérosaures et des dinosaures soient homologues mais si cela devait être le cas, cela signifierait que les ancêtres communs des ptérosaures et des dinosaures portaient des poils, et donc que les dinosaures les plus basaux furent déjà à fourrure.
Couleurs
Reconstitution des couleurs du pelage du Compsognathidae Sinosauropteryx, par Matt Martyniuk.
La découverte des dinosaures à plumes en Chine à la fin du siècle dernier et de ce siècle-ci a révolutionné nos connaissances des dinosaures et de l'évolution de leur descendants actuels les oiseaux. Mais leur découverte a permis d'amorcer une toute autre révolution dans le domaine de la paléontologie des dinosaures, celle de leur couleur. Depuis leur naissance au milieu XIX siècle jusqu'au début du XXI siècle, il était impossible de fournir des informations concrètes sur la couleur des dinosaures. En effet, les pigments de couleur ne se préservent pas à l'état fossile, et l'entièreté des fossiles de peau, de poils et de plumes des dinosaures ont perdu leur coloration à jamais. Néanmoins, grâce à un examen minutieux des poils et des plumes extrêmement bien préservés des dinosaures du Liaoning à l'aide d'un microscope électronique à balayage (SEM), les paléontologues ont découvert dans les années 2010 la présence de mélanosomes fossilisés, ces organites de cellules pigmentées comprenant la mélanine et responsables, entre autres, des couleurs arborées par les poils et les plumes. La découverte de ces mélanosomes a permis avant tout de démontrer la nature épidermique des plumes des dinosaures, qui furent vues par quelques rares paléontologues comme des fibres de collagènes.
Reconstitution des couleurs du plumage du Troodontidae Anchiornis, par Matt Martyniuk.
La combinaison de forme, taille, densité et distribution des mélanosomes a également permis de reconstituer les motifs de couleurs déployés par les plumes. Les mélanosomes qui produisent des couleurs noires et grises sont ainsi longs et étroits alors que ceux donnant des couleurs brun-rouge et marron sont courts et larges. Cette technique a jusqu'ici été appliquée à trois dinosaures, au Compsognathidae Sinosauropteryx, au Troodontidae Anchiornis et au Dromaeosauridae Microraptor. Les résultats de cette technique ont ainsi révélé, pour Sinosauropteryx, un pelage de la queue fait d'une alternance de bandes de couleurs claires et foncées, ces dernières déployant des tons plutôt brun-rouge. Chez Anchiornis, le plumage qui couvrait le corps et le cou était généralement grisé. Les ailes des membres antérieurs portaient des bandes de couleurs blanches et noires, alors que les ailes des membres postérieurs étaient blanches à leur base et noires à leur extrémité. Enfin, le plumage de la tête fut généralement grisé et tacheté de couleurs brun-rouge, tout comme les plumes de la crête qui semblent avoir été colorées plus largement de brun-rouge. Quant à Microraptor, la présence de mélanosomes de forme allongée et étroite au niveau des plumes a permis de révéler un plumage de couleur noire majoritairement iridescent.
Reconstitution hypothétique des couleurs du plumage de l'Oviraptoridae Caudipteryx, par Christophe Hendrickx.
Puisque la couleur des dinosaures fut totalement inconnue jusqu'il y a peu, et reste encore ignoré pour un grand nombre de dinosaures, les artistes de la première moitié du XX siècle dépeignaient les dinosaures dans des tons de brun, vert et gris, basant leur interprétation sur les animaux actuels comme les reptiles (crocodiles, dragons de Komodo, etc.) et les gros mammifères (éléphants, hippopotames, etc.). La palette des artistes actuels s'est néanmoins fortement élargie. Les dinosaures sont de nos jours représentés avec des couleurs vives, portant un large éventail de couleurs comme beaucoup d'oiseaux, de reptiles et de mammifères actuels, reflétant ainsi leur activité sociale complexe, une idée qui a émergé dans les années 1960.
Taille et poids
Comparaison de la taille d'Anchiornis et d'un être humain.
Si les dinosaures fascinent une grande partie du public, c'est surtout en raison de la taille gigantesque que certains d'entre eux ont atteinte. Une majorité de personnes voient d'ailleurs les dinosaures comme des animaux uniformément grands et imposants, oubliant souvent que beaucoup d'entre eux furent aussi très petits comme le montrent beaucoup de dinosaures à plumes d'une trentaine de centimètres de longueur seulement. Néanmoins, les dinosaures furent les géants de leur époque, surpassant en taille tout autre animal contemporain. Ils restent de nos jours les animaux terrestres les plus grands et les plus lourds à avoir foulé notre planète, tant au niveau des herbivores, dont les sauropodes ont les records de poids et de taille, que des carnivores, dont les plus larges dépassaient les 12 mètres de longueur.
La taille des dinosaures a fluctué durant le Trias, le Jurassique et le Crétacé; néanmoins les dinosaures ont, dès leur apparition, augmenté de taille tout au long du Mésozoïque, et atteint une taille particulièrement grande dès la fin du Trias, en témoigne le prosauropode Plateosaurus dont la taille fut estimée à 8 à 10 mètres pour un poids oscillant entre 600 à 900 kilogrammes. Cette augmentation de taille s'observe chez tous les groupes de dinosaures mis à part les sauropodes Macronaria et les théropodes coelurosaures. Ces derniers ont donné naissances aux oiseaux qui furent particulièrement petits au début de leur histoire évolutive. La réduction de taille est ainsi une tendance évolutive rare chez les dinosaures. La distribution de taille des dinosaures est également unique parmi les vertébrés, la courbe de taille des dinosaures étant positionnée au niveau d'animaux de grande taille au contraire des poissons, reptiles, mammifères et oiseaux actuels, démontrant ainsi une stratégie évolutive différente entre dinosaures et vertébrés actuels. Ceci s'observe également entre les dinosaures herbivores (sauropodomorphes et ornithopodes) et les carnivores (théropodes), ces derniers ayant autant d'individus de petite, moyenne et grande taille alors que les dinosaures herbivores comptent surtout des animaux de grande taille. Cette divergence s'explique par le fait que les dinosaures herbivores ont très vite augmenté leur taille afin d'échapper aux prédateurs et de maximiser leur efficacité digestive, au contraire des carnivores qui avaient des ressources en nourriture suffisantes parmi les dinosaures juvéniles et d'autres proies de petites tailles.
En 1990, avant la découverte d'un grand nombre de très petits dinosaures à plumes, une étude portant sur 63 genres de dinosaures évalua le poids moyen d'un dinosaure à 850 kg, un poids comparable à celui d'un grizzly, et un poids médian (qui devait osciller entre 9 kg et 5 tonnes) de près de deux tonnes, alors que le poids moyen des mammifères est de 863 grammes, soit celui d'un gros rongeur.
Plus grands et plus petits dinosaures
Les paléontologues ont de tous temps essayé d'estimer la taille et le poids des dinosaures, particulièrement des plus grands comme Brachiosaurus et des plus féroces tel Tyrannosaurus. La première méthode quantitative sérieuse fut proposée par Gregory en 1905 afin d'évaluer le poids d'Apatosaurus (alors nommé Brontosaurus). Sur base d'un modèle rigoureux de l'animal en argile et en plastique sculpté à la main, il put extrapoler le volume de l'animal en l'immergeant dans un liquide et en mesurant le volume de liquide déplacé, puis en le multipliant par la densité présumée du dinosaure (à savoir celle de l'eau) et par le facteur d'échelle du modèle. Cette technique fut suivie par d'autres auteurs jusque dans les années 1980 avant l'avènement de techniques informatiques plus robustes. Colbert, en 1962, estima ainsi le poids de Tyrannosaurus à 7 tonnes, 8,5 tonnes pour Triceratops, 28 tonnes pour Apatosaurus et 78 tonnes pour Brachiosaurus.
Seule une petite fraction des animaux morts se fossilisent, et seuls quelques spécimens découverts sont des fossiles complets, et les impressions de peaux et de tissus mous sont rares.
La reconstruction d'un squelette d'une espèce en comparant la taille et la morphologie des os avec ceux d'une autre espèce similaire mieux connue est un art inexact, et faire la recomposition des muscles et des autres organes d'un spécimen est scientifiquement difficile. On ne sera donc jamais vraiment certain de la taille des plus grands et plus petits dinosaures.
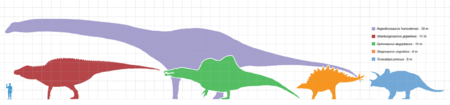
Taille des représentants les plus grands de chaque groupe de dinosaures comparée à celle d'un humain.
Parmi les dinosaures, les sauropodes étaient gigantesques, les plus grands étaient d'un ordre de grandeur plus massif que tous les animaux ayant marché depuis sur la Terre. Des mammifères préhistoriques comme l'Indricotherium et le mammouth colombien étaient des nains comparés aux sauropodes. Seule une poignée d'animaux aquatiques contemporains les approchent ou les surpassent en taille, telle la baleine bleue, qui pèse 180 tonnes et atteint 31 mètres de long au maximum.
Le plus grand et plus lourd dinosaure connu à partir de squelettes complets ou presque est le Giraffatitan (autrefois connu sous le nom de Brachiosaurus brancai). Il mesurait 12 m de haut, 22,5 m de long, et aurait pesé entre 30 et 60 tonnes (pour mémoire un éléphant de savane d'Afrique, le plus grand animal terrestre du monde, pèse en moyenne 7,7 tonnes). Le plus long dinosaure issu d'un fossile complet est le Diplodocus qui faisait 27 m (Pittsburgh, Carnegie Museum of Natural History, 1907).
Il y avait de plus grands dinosaures mais les données connues sont estimées sur quelques fossiles fragmentaires. La plupart sont des herbivores découverts dans les années 1970 ou après, parmi lesquels l'énorme Argentinosaurus, qui pourrait avoir pesé entre 80 et 100 tonnes ; le plus long de tous, le Supersaurus de 40 mètres ; et le plus grand, le Sauroposeidon de 18 mètres, qui aurait pu atteindre une fenêtre au 6 étage.
Un dinosaure encore plus grand, le Amphicoelias fragillimus, connu seulement de quelques vertèbres découvertes en 1878, pourrait avoir atteint 58 mètres de long et un poids de 120 tonnes. Le plus lourd aurait pu être le peu connu et encore débattu Bruhathkayosaurus, qui pourrait avoir atteint de 175 à 220 tonnes. On admet actuellement que Bruhathkayosaurus matleyi ne devait pas dépasser 139 tonnes, pour environ 34 mètres de long.
Le plus grand carnivore était le Spinosaurus, qui atteignait une taille de 16 à 18 mètres et pesait 9 tonnes. D'autres grands carnivores incluaient les Giganotosaurus, Mapusaurus, Tyrannosaurus rex et Carcharodontosaurus.
Sans inclure les oiseaux contemporains comme les oiseaux-mouches, les plus petits dinosaures avaient la taille d'un corbeau ou d'un poulet. Les théropodes Microraptor et Parvicursor faisaient moins de 60 cm de long.
Biologie
Comportements
Un nid de Maiasaura découvert en 1978.
L'interprétation du comportement des dinosaures non-aviens est généralement établie sur la disposition des fossiles découverts, leur habitat, les simulations par ordinateur de leurs biomécaniques (basées sur les pistes fossiles et sur des comparaisons avec des animaux actuels situés dans la même niche écologique, ainsi que l'étude des représentants actuels des dinosaures : les oiseaux). On a aussi insufflé de l'air dans les moulages de crânes de certains hadrosaures à crête pour entendre les sons que cette expérience produisit (sans que cela donne la moindre indication sur leurs rythmes et modulations). Comme telle, la compréhension actuelle du comportement des dinosaures fossiles repose donc en grande partie, comme leurs coloris, sur des spéculations, dont certaines resteront probablement controversées pendant encore longtemps. Cependant, il y a un consensus sur le fait que certaines caractéristiques qui sont communes chez les oiseaux et les crocodiles (le groupe le plus proche des dinosaures), telle que la nidification, aient été courantes chez l'ensemble des dinosaures disparus (et ce d'autant plus que des nids fossiles ont été découverts en nombre : voir plus bas).
Troupeaux
La première preuve de l'existence de troupeaux de dinosaures fut découverte en 1878 en Belgique à Bernissart. 31 Iguanodons semblaient avoir péri ensemble après être tombés dans une doline profonde et inondée ; au vu des dernières analyses, ces squelettes semblent s'être déposés lors de trois événements distincts.
D'autres sites de morts massives furent découverts. Ceux-ci, avec de nombreuses traces fossiles suggérèrent que les troupeaux ou les hordes étaient communes dans beaucoup d'espèces. Les pistes de centaines, voire de milliers d'herbivores, indiquent que les dinosaures à bec de canard pouvaient se déplacer en grands troupeaux, tel le bison ou le springbok. Des traces de sauropodes permirent de voir que ces animaux voyageaient en groupes composés de plusieurs espèces différentes, et d'autres gardaient les jeunes au milieu du troupeau afin de les protéger, d'après les traces au Davenport Ranch au Texas.
Soins parentaux
La découverte en 1978 par Jack Horner du nid du Maiasaura (« dinosaure bonne mère ») au Montana démontra que les soins parentaux duraient bien après l'éclosion chez les ornithopodes. Il y a aussi des preuves que d'autres dinosaures du Crétacé comme le sauropode Saltasaurus (découvert en 1997 en Patagonie) avaient des comportements similaires, et que ces animaux se regroupaient en immenses colonies nidificatrices comme celles des manchots.
L'Oviraptor de Mongolie a été découvert (1993) dans une position de couvaison comme celle de la poule, ce qui signifie qu'il était recouvert d'une couche de plumes isolantes qui gardait l'œuf au chaud. Des pistes fossiles ont aussi confirmé le comportement maternel parmi les sauropodes et les ornithopodes de l'île de Skye. Des nids et des œufs fossilisés ont été trouvés pour la plupart des principaux groupes de dinosaures, et il apparaît probable que les dinosaures non aviens communiquaient avec leurs petits d'une manière similaire aux dinosaures actuels (oiseaux) et aux crocodiles.
Certains dinosaures installaient leurs nids sur des sites hydrothermaux, ce qui permettait aux œufs de bénéficier d'une température idéale et régulière.
Accouplement et communication
Les crêtes de certains dinosaures, comme les marginocéphales, les théropodes et les hadrosauridae, pourraient avoir été trop fragiles pour une défense active et donc auraient probablement été utilisées pour les parades sexuelles ou à des fins d'intimidation, bien qu'il existe peu d'éléments sur le territorialisme et l'accouplement des dinosaures.
Vue d’artiste de deux Centrosaurus, ceratopsidaes herbivores de la fin du Crétacé en Amérique du Nord.
La nature des communications entre dinosaures reste également énigmatique, mais des découvertes récentes suggèrent que la crête creuse des lambeosaurines pourrait avoir fonctionné comme une caisse de résonance utilisée pour une grande variété de vocalisations.
Alimentation
Chasse
D'un point de vue comportemental, l'un des fossiles les plus importants de dinosaure a été découvert dans le désert de Gobi en 1971. Il incluait un Velociraptor attaquant un Protoceratops, prouvant physiquement que les dinosaures s'attaquaient et se mangeaient entre eux.
Cannibalisme
Bien que le cannibalisme parmi les théropodes ne soit pas une surprise, il a été confirmé par des traces de dents sur un fossile de Majungasaurus à Madagascar en 2003.
Déplacement
Basé sur les preuves fossiles existantes, il n'y avait aucune espèce de dinosaure fouisseur et peu de dinosaures grimpeurs. Puisque l'expansion des mammifères au cénozoïque vit l'apparition de nombreuses espèces fouisseuses et grimpantes, le manque de preuves pour des espèces de dinosaures similaires est quelque peu surprenant.
Une bonne compréhension de la façon dont les dinosaures se déplaçaient est la clef des modèles de comportements des espèces. La biomécanique en particulier a fourni de nombreux éléments comme la détermination de la vitesse de course des dinosaures d'après l'étude des forces exercées par leurs muscles et la gravité sur la structure de leur squelette, savoir si les diplodocides pouvaient créer un bang supersonique en balayant l'air avec leur queue en forme de fouet, déterminer si les théropodes géants devaient ralentir quand ils poursuivaient leurs proies pour éviter des blessures mortelles, et si les sauropodes pouvaient flotter.
Cycle d'activité
La théorie scientifique qui prévalait jusqu'ici est que les dinosaures avaient des modes de vie diurnes, alors que les premiers mammifères apparus étaient crépusculaires ou nocturnes.
Une étude paléontologique en 2011 de la structure oculaire (longueur de l’orbite et diamètre de l’anneau scléreux) sur 33 espèces d'archosauriens, comparée à celle de 1** espèces actuelles (dont des reptiles, des oiseaux et des mammifères), montre que les dinosaures carnivores étaient partiellement nocturnes, les dinosaures volants et ptérosaures principalement diurnes et les dinosaures herbivores surtout cathéméraux (actifs de jour comme de nuit).
Métabolisme
Une étude française sur la composition isotopique en oxygène des dents et os de 80 dinosaures du Crétacé (théropodes, sauropodes, ornithopodes et cératopsiens) provenant de gisements d'Amérique du Nord, d'Europe, d'Afrique et d'Asie, a montré que ceux-ci devaient être homéothermes. Le rapport O/O — qui dépend de la température interne de l'animal vivant — est identique à celui des mammifères et oiseaux, homéothermes, et diffère nettement de celui des reptiles actuels, ectothermes, et des chéloniens et crocodiliens fossiles du Crétacé.
La présence de structures de Havers (micro-canaux entourés d'une couche d'os concentrique au sein des squelettes) dans les os fossilisés serait également un élément en faveur du caractère endotherme.
En 2006, une étude a estimé que la température était proportionnelle à la masse et au taux de croissance, allant de 25 °C pour les petits dinosaures jusqu’à 41 °C pour les plus grands. Un modèle numérique, permettant d'estimer la température corporelle en fonction de la taille et du rythme de croissance, a été appliqué à huit espèces, du psittacosaure (Psittacosaurus mongoliensis, 12 kg) à l'apatosaure (Apatosaurus excelsus, 26 000 kg). D'après cette étude, la température interne de Sauroposeidon proteles, le plus lourd des dinosaures connus (60 tonnes), devait atteindre 48 °C. Ce modèle tendant à prouver que les gros dinosaures étaient chauffés par « homéothermie inertielle », a été remis en cause par R. Eagle et ses collègues. Ces derniers ont en effet estimé la température corporelle de grands sauropodes du Jurassique, tels Camarasaurus et Brachiosaurus, entre 36 et 38 °C, équivalente à celles de mammifères actuels.
Cependant, en 2014 une nouvelle étude comparative sur plus de 400 espèces éteintes et vivantes conclu que les dinosaures devaient avoir le sang tiède, dit mésotherme. Une étude en 2015 basée sur la température interne corrélée à la composition chimique des œufs fossilisés estime la température des sauropodes à environ 38 °C contre 32 °C pour celle des oviraptoridés.
Histoire évolutive
Origine
Une reconstitution d'Herrerasaurus, l'un des plus anciens dinosaures connus, vers -230 millions d'années au trias supérieur (vue d'artiste par Nobu Tamura).
Parmi les amniotes (eux-mêmes issus des reptiliomorphes), les dinosaures sont apparus au sein du super-ordre des archosauriens, un groupe de petits diapsides de la fin du Permien et surtout du début du Trias, jadis qualifiés de « reptiles ».
L'extinction radicale de la fin du Permien — l'extinction Permien-Trias, il y a environ 251 millions d'années, a balayé de 80 à 96 % des espèces marines, et 70 % des familles de vertébrés terrestres de l'époque, permettant à des groupes d'animaux ou de plantes de prendre leur essor par radiations évolutives, notamment les amniotes, parmi lesquels on trouve les reptiles, les ancêtres des dinosaures, et ceux des mammifères anciens et actuels. De nombreux évènements géologiques sont contemporains de cette crise.
Les petits diapsides, c'est-à-dire dont le crâne a deux paires de fosses en arrière des orbites, possèdent des os pré-dentaires à l'avant de la mandibule et des membres transversaux ou semi-dressés. Ils partent à la conquête des écosystèmes et ils divergent en trois grandes familles :
les ptérosaures jadis qualifiés de « reptiles volants » ;
les crocodiliens, dont la filiation actuelle s'est perpétuée dans un milieu amphibie ;
les dinosaures, qui, aptes à la mobilité terrestre, ont des pattes dressées dans un plan parallèle au plan sagittal de leur corps. Leurs postures redressées sont le terme d'une rapide évolution similaire à celles des mammifères, et très différentes de celle des reptiles.
L'analyse de la structure du bassin, comportant trois os pairs, ilion, pubis, ischion permet de retrouver ces différences. Ainsi le paléontologue Harry Govier Seeley a fondé dès 1887 la distinction capitale entre :
les saurischiens, littéralement « à bassin de reptiles ». Les os du bassin ont une configuration triradiée. Les oiseaux sont issus de ce groupe.
les ornithischiens, « à bassin d'oiseaux ». La ressemblance invoquée reste ici assez superficielle, mais il existe des conformations, où le pubis est parallèle à l'ischion vers le bas et l'arrière. Les oiseaux ne descendent pas de ce groupe, contrairement à ce que laisse croire ce nom.
Premiers dinosaures
Les plus anciens dinosaures connus par des fossiles sont des carnivores âgés de 225 à 230 millions d'années, Eoraptor et Herrerasaurus, découverts dans la Formation d'Ischigualasto en Argentine. Tous deux montrent déjà une certaine spécialisation, puisque ce sont des saurischiens, datant d'après la divergence entre les dinosaures ornithischiens et saurischiens. Notons que tous les dinosaures ont une denture homodonte comme les autres reptiles. Herrerasaurus est non seulement un saurischien, mais peut-être même un théropode déjà affirmé.
Des traces de pas fossiles sensiblement plus anciennes, et attribuées à des dinosaures, sont également connues. Les plus anciennes datent de 248 millions d'années et concernent « de petits reptiles quadrupèdes, de la taille d’un chat, [qui] ont laissé leurs empreintes dans l’argile il y a environ 250 millions d’années […]. Ces pas ont été découverts dans le sud-ouest [de la] Pologne, dans les Monts Sainte-Croix. […] Les empreintes […] ont été laissées il y a 248 millions d‘années par des reptiles du genre Prorotodactylus, quadrupèdes dotés de pattes à cinq doigts, dont les trois doigts centraux sont les plus fortement imprimés dans le sol. Proches les unes des autres, ces traces se distinguent de celles d’autres reptiles du groupe des crocodiles et des lézards ».
Des « traces […] plus récentes découvertes par l’équipe de Grzegorz Niedzwiedzki de l’université de Varsovie datent de 246 millions d’années. Plus grosses –une quinzaine de centimètres- elles auraient été laissées par des dinosaures du genre Sphingopus, bipèdes ».
Ces traces font probablement remonter la toute première espèce de dinosaures encore indifférenciée entre ornithischiens et saurischiens quelques millions d'années plus tôt, soit dès la fin du permien.
L'extinction du Trias-Jurassique a ensuite permis l'explosion radiative des dinosaures en libérant des niches écologiques.
Extinction
La disparition des dinosaures donna naissance à de nombreuses théories, certaines farfelues comme la destruction des dinosaures par des extraterrestres, et d'autres plus probables et scientifiquement testables. Il convient toutefois de noter que l'extinction des dinosaures est un problème sémantique : les dinosaures ne sont pas éteints puisqu'il subsiste des oiseaux.
En revanche, il y a bien eu une crise à la fin du Crétacé, il y a 65 millions d'années. Bien que cette dernière ait eu un impact moyen sur la biodiversité en général (si on la compare à la crise Permo-Triasique ou même à celle de l'Ordovicien) et qu'elle ait surtout décimé des organismes marins tels les foraminifères et non des organismes terrestres dont l'extinction relative est beaucoup moins élevée, elle est devenue très célèbre en raison de la sur-médiatisation relative aux dinosaures.
La comparaison du taux d'extinction, taxon par taxon, montre que certains des clades contemporains des dinosaures ont été très affectés (tels les plésiosaures et les ptérosaures) et d'autres beaucoup moins (crocodiliens et chéloniens par exemple). Cette crise a éliminé les dinosaures non-aviens vivant à cette époque.
Les causes les plus probables ayant induit la crise K-T sont :
la chute d'un astéroïde ou d'une comète d'une dizaine de kilomètres de diamètre provoquant une catastrophe majeure qui, par le biais de débris dus à la collision, plongea la Terre dans l'obscurité et le froid pendant plusieurs années, empêchant ainsi la photosynthèse, ce qui induisit un appauvrissement massif en plantes et surtout en plancton et conduisit à l'extinction de nombreuses espèces dépendant de ces ressources quel que soit leur niveau trophique. Cette théorie, de nos jours très argumentée (cratère de Chicxulub au Mexique daté de –65 Ma), permet notamment d'expliquer dans une certaine mesure les survies différentielles des taxons. De plus, à la suite de découvertes récentes (avril 2008) de cénosphères (sphères microscopiques se formant à la suite de la combustion de charbon et de pétrole brut) dans les sédiments de la crise K-T, les chercheurs pensent que la météorite aurait pu s'écraser dans une réserve pétrolière, et projeter dans l’atmosphère de grandes quantités d’hydrocarbures enflammés expliquant la disparition de la majorité des espèces. Une étude publiée le 7 février 2013 dans la revue Science et s'appuyant sur des techniques de datation radiométriques de haute précision, indique que la météorite se serait écrasée sur Terre il y a 66 038 000 d'années, soit au maximum 32 000 ans avant l'extinction des dinosaures, ce qui renforcerait d'autant la probabilité d'un lien de causalité entre les deux évènements.
des éruptions à la fin du Crétacé d'un "supervolcan", lié à la présence d'un point chaud placé actuellement sous l'île de La Réunion et qui forma les trapps du Deccan, en Inde pendant une période assez courte au niveau des temps géologiques (au moins 1 million d'années), provoquant un cataclysme planétaire en modifiant le climat et réduisant la photosynthèse en plongeant la Terre dans une pénombre par le biais de nuages de cendres et de brume sulfureuse. Cette théorie est aussi argumentée que la théorie précédente, entre autres par la datation de –65 Ma des trapps du Deccan, dont la superficie équivaut à plusieurs fois la France, et par des résultats sur la baisse de la luminosité à la surface de la Terre lors de l'éruption récente du Pinatubo (avril 1991) aux Philippines. Cette théorie recoupe en partie celle de la collision météorique sur les effets atmosphériques.
des régressions marines très importantes à la fin du Crétacé changeant la conformation et l'étendue des milieux littoraux et benthiques tout en induisant un important changement climatique. Cette théorie est elle aussi basée sur des preuves solides et on sait maintenant qu'une très grande régression marine a eu lieu au crétacé supérieur.
Ces trois théories sont basées sur des faits et la crise K-T pourrait être la conséquence de la quasi-simultanéité de ces trois événements. Les avis divergent en ce qui concerne l'importance relative de chacun d'eux.
Dans une étude publiée le 18 mars 2016 dans le Proceedings of the National Academy of Sciences (PNAS), plusieurs paléontologues de l'Université de Reading ont révélé que 40 millions d'années avant leur extinction, les dinosaures avaient déjà commencé à disparaître; l'hypothèse à privilégier serait alors celle d'une disparition progressive provoquée par la séparation des continents et une forte activité volcanique.
Représentants actuels

Archaeopteryx, Berlin, 1863
Le premier fossile d'oiseau, l'Archéoptéryx, du Jurassique supérieur, a été découvert en Bavière en 1861. Sa grande ressemblance avec certains petits dinosaures carnivores bipèdes, comme les Compsognathus, a immédiatement fait apparaître la théorie selon laquelle les oiseaux descendraient d'un groupe de dinosaures au sein des Cœlurosauriens.
Pendant un siècle, cette théorie est restée très controversée, voire rejetée. En effet, les oiseaux ont des clavicules, alors que les cœlurosauriens n'en avaient pas. Mais l'importance accordée aux clavicules est excessive: les carnivores n'ont pas de clavicules; ils n'en sont pas moins des mammifères.
Depuis les années 1970, cependant, des Cœlurosauriens dotés de clavicules ont été découverts (et même dans des groupes moins dérivés) et la théorie dinosaurienne sur l'origine des oiseaux est redevenue dominante. Dans les années 1990, de nombreux fossiles de dinosaures à plumes ont été découverts, principalement dans la région du Liaoning, en Chine et ont contribué à réactiver cette théorie. Il s'agit dans certains cas d'oiseaux primitifs, et dans d'autres de dinosaures non aviens à plumes ou proto-plumes. L'interprétation qui est faite de ces découvertes est qu'une ou plusieurs espèces de dinosaure cœlurosaurien (voire l'ancêtre des Cœlurosauriens lui-même) a développé le caractère « plume », semble-t-il sans l'utiliser pour le vol, et a donné naissance à des espèces assez nombreuses de dinosaures à plumes. Une de ces espèces de dinosaures à plume aurait, des millions d'années plus tard, donné naissance à l'ancêtre commun à tous les oiseaux. En 2011, au Canada, onze plumes de dinosaures, colorées (du marron au noir) ont été retrouvées dans de l'ambre, datant de 78 à 79 millions d'années (fin crétacé) avec barbes et barbules visibles, semblables à ceux d'oiseaux contemporains, avec des indices d'imperméabilité pouvant laisser penser qu'il s'agissait (dans ce cas) d'animaux semi-aquatiques. Divers indices laissent penser que les plumes de dinosaures étaient souvent colorées.
Bien que cette théorie faisant des oiseaux un sous-groupe des dinosaures soit très largement dominante, il reste des contestataires, regroupés sous l'acronyme « BAND » : Birds Are Not Dinosaurs.
« Reptiles » contemporains des dinosaures
De nombreuses lignées jadis qualifiées de « reptiliennes » vivaient aux mêmes périodes géologiques que les dinosaures, et ont parfois été confondues avec les dinosaures par le cinéma ou la littérature. Les plus connues sont :
les ptérosaures (qualifiés jadis de « reptiles volants ») incluant le ptéranodon et le ptérodactyle ;
les plésiosaures, les pliosaures et les ichthyosaures (qualifiés jadis de « reptiles marins »), ainsi que les mosasaures (les seuls que l'on pourrait dénommer « reptiles marins » puisque ce sont des lépidosaures, comme les actuels varans).
Aucune de ces lignées n'est scientifiquement classée comme « dinosaurienne ».
Histoire des découvertes
Les fossiles de grands animaux mésozoïques sont connus depuis l'aube de l'humanité, mais leurs véritables identifications n'ont émergé qu'après des millénaires d'interprétations mythologiques, et après plusieurs décennies de postulats fantaisistes même au sein d'une géologie et d'une paléontologie scientifiques, mais manquant, aux débuts, d'un nombre suffisant de données.
Pour les Chinois c'étaient des os de dragons, pour les Européens des restes des Géants bibliques et d'autres créatures tuées par le Déluge. Georges Cuvier décrit un lézard marin, le mosasaure (contemporain des dinosaures, mais qui n'en était pas un), dès 1808. Les premières espèces identifiées et baptisées sont l’iguanodon, découvert en 1822 par le géologue anglais Gideon Mantell, qui a remarqué des similitudes entre ses fossiles et les os de l’iguane contemporain. Le premier article scientifique sur les dinosaures paraît deux ans plus tard. Il est publié par le révérend William Buckland, professeur de géologie à l'université d’Oxford, et concerne Megalosaurus bucklandii, dont un fossile avait été découvert près d’Oxford. L'étude de ces « grands lézards fossiles » fait l'objet d'un grand intérêt dans les cercles scientifiques européens et américains, et le paléontologue anglais Richard Owen invente le terme « dinosaure » en 1842. Il remarque que les restes précédemment trouvés (Iguanodon, Megalosaurus et Hylaeosaurus) ont de nombreux caractères en commun, et décide de créer un nouveau groupe taxonomique. Avec l'aide du prince Albert de Saxe-Cobourg-Gotha, mari de la reine Victoria, il crée le Muséum d'histoire naturelle de Londres, à South Kensington (Natural History Museum), pour exposer la collection nationale de fossiles de dinosaures, ainsi que quelques autres objets d'intérêt botanique et géologique.
Vulgarisateur bénéficiant de gros budgets pour faire voir et faire toucher ce qu'il croit être une reconstitution exacte, Richard Owen rend populaire sa dénomination qui atteint un apogée dans les foules curieuses vers 1890. Le naturaliste Thomas Henry Huxley propose ornithoscélidés, littéralement « à la cuisse d'oiseaux ». Hermann de Meyer préfère pachypodes c'est-à-dire « aux pieds épais ». Ces dénominations ne quittent pas le cercle de lecteurs savants. Mais, après 1887, les spécialistes de sciences naturelles lassés du spectacle d'Owen, ordonnent avec rigueur les différentes familles de Saurischiens et d'Ornithischiens.
En 1858 le premier fossile de dinosaure américain est découvert, dans des marnières près de la petite ville de Haddonfield, dans le New Jersey (ce n'est pas le premier fossile de dinosaure trouvé en Amérique, mais le premier identifié comme tel). L’animal est nommé Hadrosaurus foulkii, du nom de la ville et de son découvreur : William Parker Foulke. Cette découverte est capitale car il s'agissait du premier squelette presque complet découvert, et Joseph Leidy met en évidence sans doute possible que l'animal était bipède. Jusqu’alors la plupart des scientifiques croyaient que les dinosaures marchaient à quatre pattes comme les lézards.
La découverte de l'Hadrosaurus bipède marque le début effervescent d'une chasse aux fossiles de dinosaures aux États-Unis. La lutte acharnée entre Edward Drinker Cope, amateur fortuné de géologie et de paléontologie, natif de Philadelphie et Othniel Charles Marsh, heureux new-yorkais héritier de Georges Peabody à 21 ans et prospecteur infatigable, est connue sous le nom de « guerre des os » (Bone Wars). Leur querelle irrémédiable après 1870, née d'une rivalité dans la chasse aux dinosaures démultipliée par de nombreuses brouilles dont Como Bluff, gisement mis au jour par deux ouvriers de l'Union Pacific et finalement exploité par O.C. Marsh, dure presque 30 ans, et finit en 1879 quand Cope mourut après avoir dépensé toute sa fortune dans cette quête. Marsh qui lui survécut vingt années sortit vainqueur, mais n'évita la ruine que grâce à l’aide financière inespérée de l’Organisation Géologique des États-Unis (United States Geological Survey). La collection d'Edward Cope, auteur prolifique de plus de 1800 articles scientifiques depuis ses dix-huit ans, ce qui ne l'empêcha pas de placer la tête sur la queue d'un plésiosaure, se trouve aujourd'hui au muséum d'histoire naturelle de New York, celle de Marsh au muséum d'histoire naturelle de Peabody, à l'université Yale.
L'assistant de Cope, Charles Hazelius Sternberg aidé de ses trois fils, reprend le flambeau en entreprenant, au début du siècle suivant, des missions en Alberta, au Kansas, dans le Montana et le Wyoming. Dans ce dernier État, il relève en 1908 la première empreinte de peau momifiée de ce qui s'avère être un Anatosaurus. Une seconde découverte similaire est faite en 1910. Une partie de son équipe rejoint en 1913 Barnum Brown, qui, parfois avec son ami Henry Fairfield Osborn, prospecte depuis 1910 les berges de la Red Deer river entre l'Alberta et le Montana. Les barges des chercheurs, perturbé dans leur repérage au fil de l'eau par d'agaçants moustiques, dérivent avec le courant. Mais la moisson de trouvailles en vaut la peine : squelettes de théropodes, aratopsiens, hadrosaures...
Les chercheurs du II Reich annoncent la généralisation de la chasse lointaine et systématique, par leurs succès dans les colonies allemandes d'Afrique occidentale et orientale. Dans l'actuelle Tanzanie, le site de Tendaguru livre des grands sauropodes de la famille des Brachiosauridés et des petits stégosauriens genre Kentrosaurus aux paléontologues allemands. Werner Janensch supervise la collection rapatriée au musée d'histoire naturelle de Berlin. Edwin Hennig poursuit les investigations en Afrique entre 1908 et 1912.
C'est grâce à une exploration asiatique à la quête de l'origine de l'homme que la Smithsonian Institution, par l'intermédiaire de son musée d'histoire naturelle, cumule les trouvailles décisives dans le désert de Gobi entre 1922 et 1925. La mission de l'American Museum of Natural History, commandé par Roy Chapman Andrews, animée par Osborn et Granger, découvre des œufs et des nids de Protoceratops andrewsi.
La Chine et ses confins avaient déjà attiré une expédition russe entre 1915 et 1917. Les missions russes, source de prestige, reprennent en 1946 et 1948. Les Chinois rentrent tôt dans la course et lancent leurs propres missions scientifiques après 1933. Après le refroidissement des relations sino-soviétiques, elles se cantonnent à la Mongolie. Les missions russo-mongoles et polono-mongoles se succèdent dans les années 1960 et 1970. Le Gobi est alors l'éden des chasseurs de dinosaures. Plus que les rares revues spécialisées, journaux et radio relatent les péripéties aventureuses.
La Chine investit avec force la spécialité de la paléontologie des vertébrés. Le temps du terrible dragon Kong-Hong reconnu dans les entrailles de la terre et vénéré laisse la place à l'analyse scientifique. Des spécialistes de renom émergent tels C.C. Young alias Yang Zhoug-jian et le professeur Dong Zhimming qui découvrent plus de cent espèces inconnues en Chine: Sauropode Mamenchisaurus, Hadrosaure Tsintaosaurus, Prosauropode Lufengosaurus, Stegosaure Tuojiangosaurus…
La connaissance de la faune du Mésozoïque s'accroît aussi avec l'expédition franco-norvégienne du Spitzberg en 1968, les paléontologues français y sont avantagés par la connaissance préalable de cette période géologique au Sahara et en Afrique du Nord.
La recherche de fossiles s'est étendue à toute la surface du globe, y compris en Antarctique, où un ankylosaure, Antarctopelta, est découvert en 1986 sur l'île James-Ross, mais dont la description n'intervient que vingt ans après. C'est cependant en 1994 qu'un dinosaure habitant réellement l'Antarctique, Cryolophosaurus ellioti, est décrit dans un journal scientifique. Les zones particulièrement intéressantes sont aujourd'hui l'Amérique du Sud, et surtout l'Argentine et la Chine, dont le sous-sol a révélé de nombreux squelettes très bien conservés.
Classification
Il n'y a pas encore si longtemps, on classait dans les dinosaures : les ichthyosaures, les pliosaures, les plésiosaures (trois groupes de reptiles aquatiques) et les ptérosaures (reptiles volants) qui, aujourd'hui, sont considérés comme des lignées évolutives indépendantes des dinosaures parmi les reptiles ayant vécu au Mésozoïque. Les dinosaures étaient un groupe d'animaux extrêmement divers ; selon une étude de 2006, 527 genres de dinosaures ont été décrits avec certitude, et plus de 1 844 genres sont encore à classer. Certains d'entre eux étaient herbivores, d'autres carnivores. Certains dinosaures étaient bipèdes, d'autres étaient quadrupèdes et certains, tels Ammosaurus et Iguanodon, pouvaient marcher aussi bien sur deux ou quatre pattes.
Les dinosaures sont nommés de façon précise selon leur genre et leur espèce. Souvent, ce nom d'espèce est donné en fonction du nom du lieu de sa découverte comme Iguanodo bernissartensis du nom de la localité de Bernissart, en Belgique, où fut découvert un troupeau de 31 iguanodons, ou bien Saltasaurus, le dinosaure de la rivière Salta en Argentine. Une particularité anatomique peut servir de base pour baptiser une espèce, comme c'est le cas pour le Triceratops, la face à trois cornes. Ou c'est le nom d'un paléontologue connu qui est utilisé, comme pour Othnielia du nom d'Othniel Charles Marsh. Avec l'essor de la paléontologie, les noms d'espèces s'internationalisent. Les racines grecques et latines sont remplacées parfois par des racines chinoises, mongoles ou africaines comme Nqwebasaurus venant du xhosa, une langue d'Afrique du Sud.
Tableau récapitulatif
Sur la base du code international de nomenclature zoologique, on peut reconnaitre de 630 à 650 genres de dinosaures, rassemblant plusieurs milliers d'espèces de dinosaures (une trentaine de nouvelles espèces sont décrites tous les ans). Un peu plus de la moitié de ces espèces ne sont représentées que par un seul spécimen (souvent incomplet) et moins de 20 % des espèces sont connues par plus de cinq spécimens. Ce tableau général fournit des liens entre les principaux groupes de dinosaures. On trouvera à droite quelques genres illustrant les ordres et les familles représentés.
Saurischia Herrerasauridae Herrerasaurus Staurikosaurus Theropoda Coelophysoidea Coelophysis Liliensternus Dilophosaurus Neotheropoda Ceratosauria Ceratosaurus Majungasaurus Carnotaurus Tetanurae Megalosauroidea Spinosaurus Suchomimus Torvosaurus Avetheropoda Allosaurus Tyrannosaurus Velociraptor Sauropodomorpha Sauropodomorphes basaux Anchisaurus Plateosaurus Massospondylus Sauropoda Sauropodes basaux Mamenchisaurus Shunosaurus Omeisaurus Neosauropoda Diplodocoidea Diplodocus Apatosaurus Amargasaurus Macronaria Brachiosaurus Camarasaurus Ampelosaurus Ornithischia Heterodontosauridae Heterodontosaurus Abrictosaurus Genasauria Thyreophora Thyréophores basaux Scelidosaurus Scutellosaurus Eurypoda Stegosauria Stegosaurus Kentrosaurus Huayangosaurus Ankylosauria Euoplocephalus Edmontonia Ankylosaurus Cerapoda Marginocephalia Pachycephalosauria Prenocephale Pachycephalosaurus Stegoceras Ceratopsia Triceratops Protoceratops Torosaurus Ornithopoda Ornithopodes basaux Hypsilophodon Agilisaurus Leællynasaura Iguanodontia Iguanodon Edmontosaurus Parasaurolophus
Voir aussi liste des dinosaures.
Infra-ordre des Coelophysoidea
Infra-ordre des Ceratosauria Famille des Ceratosauridae Super-famille des Abelisauroidea Famille des Noasauridae Famille des Abelisauridae
Famille des Ceratosauridae
Super-famille des Abelisauroidea Famille des Noasauridae Famille des Abelisauridae
Famille des Noasauridae
Famille des Abelisauridae
Infra-ordre des Tetanurae Super-famille des Megalosauroidea Famille des Piatnitzkysauridae Famille des Megalosauridae Famille des Spinosauridae Sous-famille des Baryonychinae Sous-famille des Spinosaurinae Super-famille des Neotetanurae Division des Allosauroidea Famille des Sinraptoridae Famille des Allosauridae Division des Carcharodontosauria Famille des Carcharodontosauridae Famille des Neovenatoridae Division des Coelurosauria Super-famille des Tyrannosauroidea Famille des Proceratosauridae Famille des Megaraptoridae Famille des Tyrannosauridae Sous-famille des Tyrannosaurinae Famille des Compsognathidae Sous-division des Maniraptoriformes Infradivision des Ornithomimosauria Famille des Deinocheiridae Famille des Ornithomimidae Infradivision des Maniraptora Infradivision des Therizinosauria Super-famille des Therizinosauroidea Famille des Therizinosauridae Super-famille des Alvarezsauroidea Infra-ordre des Oviraptorosauria Famille des Caudipteridae Super-famille des Caenagnathoidea Famille des Caenagnathidae Famille des Oviraptoridae Paraves Infra-ordre des Deinonychosauria Famille des Dromaeosauridae Famille des Troodontidae Avialae
Super-famille des Megalosauroidea Famille des Piatnitzkysauridae Famille des Megalosauridae Famille des Spinosauridae Sous-famille des Baryonychinae Sous-famille des Spinosaurinae
Famille des Piatnitzkysauridae
Famille des Megalosauridae
Famille des Spinosauridae Sous-famille des Baryonychinae Sous-famille des Spinosaurinae
Sous-famille des Baryonychinae
Sous-famille des Spinosaurinae
Super-famille des Neotetanurae Division des Allosauroidea Famille des Sinraptoridae Famille des Allosauridae Division des Carcharodontosauria Famille des Carcharodontosauridae Famille des Neovenatoridae Division des Coelurosauria Super-famille des Tyrannosauroidea Famille des Proceratosauridae Famille des Megaraptoridae Famille des Tyrannosauridae Sous-famille des Tyrannosaurinae Famille des Compsognathidae Sous-division des Maniraptoriformes Infradivision des Ornithomimosauria Famille des Deinocheiridae Famille des Ornithomimidae Infradivision des Maniraptora Infradivision des Therizinosauria Super-famille des Therizinosauroidea Famille des Therizinosauridae Super-famille des Alvarezsauroidea Infra-ordre des Oviraptorosauria Famille des Caudipteridae Super-famille des Caenagnathoidea Famille des Caenagnathidae Famille des Oviraptoridae Paraves Infra-ordre des Deinonychosauria Famille des Dromaeosauridae Famille des Troodontidae Avialae
Division des Allosauroidea Famille des Sinraptoridae Famille des Allosauridae Division des Carcharodontosauria Famille des Carcharodontosauridae Famille des Neovenatoridae
Famille des Sinraptoridae
Famille des Allosauridae Division des Carcharodontosauria Famille des Carcharodontosauridae Famille des Neovenatoridae
Division des Carcharodontosauria Famille des Carcharodontosauridae Famille des Neovenatoridae
Famille des Carcharodontosauridae
Famille des Neovenatoridae
Division des Coelurosauria Super-famille des Tyrannosauroidea Famille des Proceratosauridae Famille des Megaraptoridae Famille des Tyrannosauridae Sous-famille des Tyrannosaurinae Famille des Compsognathidae Sous-division des Maniraptoriformes Infradivision des Ornithomimosauria Famille des Deinocheiridae Famille des Ornithomimidae Infradivision des Maniraptora Infradivision des Therizinosauria Super-famille des Therizinosauroidea Famille des Therizinosauridae Super-famille des Alvarezsauroidea Infra-ordre des Oviraptorosauria Famille des Caudipteridae Super-famille des Caenagnathoidea Famille des Caenagnathidae Famille des Oviraptoridae Paraves Infra-ordre des Deinonychosauria Famille des Dromaeosauridae Famille des Troodontidae Avialae
Super-famille des Tyrannosauroidea Famille des Proceratosauridae Famille des Megaraptoridae Famille des Tyrannosauridae Sous-famille des Tyrannosaurinae
Famille des Proceratosauridae
Famille des Megaraptoridae
Famille des Tyrannosauridae Sous-famille des Tyrannosaurinae
Sous-famille des Tyrannosaurinae
Famille des Compsognathidae
Sous-division des Maniraptoriformes Infradivision des Ornithomimosauria Famille des Deinocheiridae Famille des Ornithomimidae Infradivision des Maniraptora Infradivision des Therizinosauria Super-famille des Therizinosauroidea Famille des Therizinosauridae Super-famille des Alvarezsauroidea Infra-ordre des Oviraptorosauria Famille des Caudipteridae Super-famille des Caenagnathoidea Famille des Caenagnathidae Famille des Oviraptoridae Paraves Infra-ordre des Deinonychosauria Famille des Dromaeosauridae Famille des Troodontidae Avialae
Infradivision des Ornithomimosauria Famille des Deinocheiridae Famille des Ornithomimidae
Famille des Deinocheiridae
Famille des Ornithomimidae
Infradivision des Maniraptora Infradivision des Therizinosauria Super-famille des Therizinosauroidea Famille des Therizinosauridae Super-famille des Alvarezsauroidea Infra-ordre des Oviraptorosauria Famille des Caudipteridae Super-famille des Caenagnathoidea Famille des Caenagnathidae Famille des Oviraptoridae Paraves Infra-ordre des Deinonychosauria Famille des Dromaeosauridae Famille des Troodontidae Avialae
Infradivision des Therizinosauria Super-famille des Therizinosauroidea Famille des Therizinosauridae
Super-famille des Therizinosauroidea Famille des Therizinosauridae
Famille des Therizinosauridae
Super-famille des Alvarezsauroidea
Infra-ordre des Oviraptorosauria Famille des Caudipteridae Super-famille des Caenagnathoidea Famille des Caenagnathidae Famille des Oviraptoridae
Famille des Caudipteridae
Super-famille des Caenagnathoidea Famille des Caenagnathidae Famille des Oviraptoridae
Famille des Caenagnathidae
Famille des Oviraptoridae
Paraves Infra-ordre des Deinonychosauria Famille des Dromaeosauridae Famille des Troodontidae Avialae
Infra-ordre des Deinonychosauria Famille des Dromaeosauridae Famille des Troodontidae
Famille des Dromaeosauridae
Famille des Troodontidae
Avialae
Sauropodomorpha basaux
Anchisauria (en) Infra-ordre des Prosauropoda Famille des Anchisauridae Plateosauria Famille des Plateosauridae Famille des Massospondylidae Infra-ordre des Sauropoda Sauropoda basaux Famille des Melanorosauridae Eusauropoda Eusauropoda basaux Famille des Mamenchisauridae Famille des Cetiosauridae Turiasauria Division des Neosauropoda Super-famille des Diplodocoidea Famille des Rebbachisauridae Flagellicaudata Famille des Dicraeosauridae Famille des Diplodocidae Sous-division des Macronaria Macronaria basaux Camarasauromorpha Famille des Camarasauridae Infra-division des Titanosauriformes Famille des Brachiosauridae Somphospondyli Somphospondyli basaux Super-famille des Titanosauria Titanosauria basaux Lithostrotia Lognkosauria Famille des Nemegtosauridae Famille des Saltasauridae
Infra-ordre des Prosauropoda Famille des Anchisauridae Plateosauria Famille des Plateosauridae Famille des Massospondylidae
Famille des Anchisauridae
Plateosauria Famille des Plateosauridae Famille des Massospondylidae
Famille des Plateosauridae
Famille des Massospondylidae
Infra-ordre des Sauropoda Sauropoda basaux Famille des Melanorosauridae Eusauropoda Eusauropoda basaux Famille des Mamenchisauridae Famille des Cetiosauridae Turiasauria Division des Neosauropoda Super-famille des Diplodocoidea Famille des Rebbachisauridae Flagellicaudata Famille des Dicraeosauridae Famille des Diplodocidae Sous-division des Macronaria Macronaria basaux Camarasauromorpha Famille des Camarasauridae Infra-division des Titanosauriformes Famille des Brachiosauridae Somphospondyli Somphospondyli basaux Super-famille des Titanosauria Titanosauria basaux Lithostrotia Lognkosauria Famille des Nemegtosauridae Famille des Saltasauridae
Sauropoda basaux
Famille des Melanorosauridae Eusauropoda Eusauropoda basaux Famille des Mamenchisauridae Famille des Cetiosauridae Turiasauria Division des Neosauropoda Super-famille des Diplodocoidea Famille des Rebbachisauridae Flagellicaudata Famille des Dicraeosauridae Famille des Diplodocidae Sous-division des Macronaria Macronaria basaux Camarasauromorpha Famille des Camarasauridae Infra-division des Titanosauriformes Famille des Brachiosauridae Somphospondyli Somphospondyli basaux Super-famille des Titanosauria Titanosauria basaux Lithostrotia Lognkosauria Famille des Nemegtosauridae Famille des Saltasauridae
Eusauropoda Eusauropoda basaux Famille des Mamenchisauridae Famille des Cetiosauridae Turiasauria Division des Neosauropoda Super-famille des Diplodocoidea Famille des Rebbachisauridae Flagellicaudata Famille des Dicraeosauridae Famille des Diplodocidae Sous-division des Macronaria Macronaria basaux Camarasauromorpha Famille des Camarasauridae Infra-division des Titanosauriformes Famille des Brachiosauridae Somphospondyli Somphospondyli basaux Super-famille des Titanosauria Titanosauria basaux Lithostrotia Lognkosauria Famille des Nemegtosauridae Famille des Saltasauridae
Eusauropoda basaux
Famille des Mamenchisauridae
Famille des Cetiosauridae
Turiasauria
Division des Neosauropoda Super-famille des Diplodocoidea Famille des Rebbachisauridae Flagellicaudata Famille des Dicraeosauridae Famille des Diplodocidae Sous-division des Macronaria Macronaria basaux Camarasauromorpha Famille des Camarasauridae Infra-division des Titanosauriformes Famille des Brachiosauridae Somphospondyli Somphospondyli basaux Super-famille des Titanosauria Titanosauria basaux Lithostrotia Lognkosauria Famille des Nemegtosauridae Famille des Saltasauridae
Super-famille des Diplodocoidea Famille des Rebbachisauridae Flagellicaudata Famille des Dicraeosauridae Famille des Diplodocidae
Famille des Rebbachisauridae
Flagellicaudata Famille des Dicraeosauridae Famille des Diplodocidae
Famille des Dicraeosauridae
Famille des Diplodocidae
Sous-division des Macronaria Macronaria basaux Camarasauromorpha Famille des Camarasauridae Infra-division des Titanosauriformes Famille des Brachiosauridae Somphospondyli Somphospondyli basaux Super-famille des Titanosauria Titanosauria basaux Lithostrotia Lognkosauria Famille des Nemegtosauridae Famille des Saltasauridae
Macronaria basaux
Camarasauromorpha Famille des Camarasauridae Infra-division des Titanosauriformes Famille des Brachiosauridae Somphospondyli Somphospondyli basaux Super-famille des Titanosauria Titanosauria basaux Lithostrotia Lognkosauria Famille des Nemegtosauridae Famille des Saltasauridae
Famille des Camarasauridae
Infra-division des Titanosauriformes Famille des Brachiosauridae Somphospondyli Somphospondyli basaux Super-famille des Titanosauria Titanosauria basaux Lithostrotia Lognkosauria Famille des Nemegtosauridae Famille des Saltasauridae
Famille des Brachiosauridae
Somphospondyli Somphospondyli basaux Super-famille des Titanosauria Titanosauria basaux Lithostrotia Lognkosauria Famille des Nemegtosauridae Famille des Saltasauridae
Somphospondyli basaux
Super-famille des Titanosauria Titanosauria basaux Lithostrotia Lognkosauria Famille des Nemegtosauridae Famille des Saltasauridae
Titanosauria basaux
Lithostrotia Lognkosauria Famille des Nemegtosauridae Famille des Saltasauridae
Lognkosauria
Famille des Nemegtosauridae
Famille des Saltasauridae
Thyreophora basaux
Thyreophoroidea Thyreophoroidea basaux Infra-ordre des Stegosauria Famille des Stegosauridae Sous-famille des Stegosaurinae Famille des Huayangosauridae Infra-ordre des Ankylosauria Famille des Nodosauridae Famille des Ankylosauridae Sous-famille des Ankylosaurinae
Thyreophoroidea basaux
Infra-ordre des Stegosauria Famille des Stegosauridae Sous-famille des Stegosaurinae Famille des Huayangosauridae
Famille des Stegosauridae Sous-famille des Stegosaurinae
Sous-famille des Stegosaurinae
Famille des Huayangosauridae
Infra-ordre des Ankylosauria Famille des Nodosauridae Famille des Ankylosauridae Sous-famille des Ankylosaurinae
Famille des Nodosauridae
Famille des Ankylosauridae Sous-famille des Ankylosaurinae
Sous-famille des Ankylosaurinae
Infra-ordre des Pachycephalosauria Pachycephalosauria basaux Pachycephalosauridae Pachycephalosauridae basaux Sous-famille des Pachycephalosaurinae
Pachycephalosauria basaux Pachycephalosauridae Pachycephalosauridae basaux Sous-famille des Pachycephalosaurinae
Pachycephalosauridae Pachycephalosauridae basaux Sous-famille des Pachycephalosaurinae
Pachycephalosauridae basaux Sous-famille des Pachycephalosaurinae
Sous-famille des Pachycephalosaurinae
Infra-ordre des Ceratopsia Famille des Psittacosauridae Neoceratopsia Neoceratopsia basaux Coronosauria Famille des Leptoceratopsidae Famille des Protoceratopsidae Famille des Ceratopsidae Sous-famille des Centrosaurinae Sous-famille des Chasmosaurinae
Famille des Psittacosauridae
Neoceratopsia Neoceratopsia basaux Coronosauria Famille des Leptoceratopsidae Famille des Protoceratopsidae Famille des Ceratopsidae Sous-famille des Centrosaurinae Sous-famille des Chasmosaurinae
Neoceratopsia basaux Coronosauria Famille des Leptoceratopsidae Famille des Protoceratopsidae Famille des Ceratopsidae Sous-famille des Centrosaurinae Sous-famille des Chasmosaurinae
Coronosauria
Famille des Leptoceratopsidae
Famille des Protoceratopsidae
Famille des Ceratopsidae Sous-famille des Centrosaurinae Sous-famille des Chasmosaurinae
Sous-famille des Centrosaurinae
Sous-famille des Chasmosaurinae
Ornithopoda basaux
Iguanodontia Iguanodontia basaux Dryomorpha Dryomorpha basaux Ankylopollexia Ankylopollexia basaux Super-famille des Iguanodontoidea Iguanodontoidea basaux Famille des Hadrosauridae Sous-famille des Hadrosaurinae Sous-famille des Lambeosaurinae
Iguanodontia basaux
Dryomorpha Dryomorpha basaux Ankylopollexia Ankylopollexia basaux Super-famille des Iguanodontoidea Iguanodontoidea basaux Famille des Hadrosauridae Sous-famille des Hadrosaurinae Sous-famille des Lambeosaurinae
Dryomorpha basaux
Ankylopollexia Ankylopollexia basaux Super-famille des Iguanodontoidea Iguanodontoidea basaux Famille des Hadrosauridae Sous-famille des Hadrosaurinae Sous-famille des Lambeosaurinae
Ankylopollexia basaux
Super-famille des Iguanodontoidea Iguanodontoidea basaux Famille des Hadrosauridae Sous-famille des Hadrosaurinae Sous-famille des Lambeosaurinae
Iguanodontoidea basaux
Famille des Hadrosauridae Sous-famille des Hadrosaurinae Sous-famille des Lambeosaurinae
Sous-famille des Hadrosaurinae
Sous-famille des Lambeosaurinae
Cladogramme du clade des Dinosauria ─o Dinosauria
│
├─o Ornithischia
│ ├─o Lesothosaurus
│ └─o Genasauria
│ │
│ ├─o Thyreophora
│ │ ├─o Scutellosaurus
│ │ └─o
│ │ ├─o Scelidosaurus
│ │ └─o Eurypoda
│ │ ├─o Stegosauria
│ │ └─o Ankylosauria
│ │
│ └─o Neornithischia
│ ├─o Marginocephalia
│ │ ├─o Pachycephalosauria
│ │ └─o Ceratopsia
│ └─o Ornithopoda
│
└─o Saurischia
│
├─o Sauropodomorpha
│ ├─o Prosauropoda (paraphylétique)
│ └─o Sauropoda
│
└─o Theropoda
├─o Herrerasauridae
└─o Neotheropoda
└─o Coelophysoidea
└─o Averostra
├─o Ceratosauria
│ ├─o Ceratosauridae
│ └─o Abelisauroidea
│ ├─o Noasauridae
│ └─o Abelisauridae
└─o Tetanurae
├─o Megalosauroidea
│ ├─o Piatnitzkysauridae
│ └─o Megalosauria
│ ├─o Megalosauridae
│ └─o Spinosauridae
└─o Avetheropoda
├─o Allosauroidea
│ ├─o Metriacanthosauridae
│ └─o Allosauria
│ ├─o Allosauridae
│ └─o Carcharodontosauria
│ ├─o Neovenatoridae
│ └─o Carcharodontosauridae
└─o Coelurosauria
├─o Tyrannosauroidea
│ ├─o Proceratosauridae
│ └─o
│ ├─o Megaraptora
│ └─o Tyrannosauridae
└─o
├─o Compsognathidae
└─o Maniraptoriformes
├─o Ornithomimosauria
│ ├─o Deinocheiridae
│ └─o Ornithomimidae
└─o Maniraptora
├─o Alvarezsauroidea
│ └─o Alvarezsauridae
└─o
├─o Therizinosauria
│ └─o Therzinosauroidea
│ └─o Therizinosauridae
└─o Pennaraptora
├─o Oviraptorosauria
│ ├─o Caudipteridae
│ └─o Caenagnathoidea
│ ├─o Caenagnathidae
│ └─o Oviraptoridae
└─o Paraves
├─o Dromaeosauridae
│ ├─o Unenlagiinae
│ └─o
│ ├─o Microraptorinae
│ └─o Eudromaeosauria
│ ├─o Dromaeosaurinae
│ └─o Velociraptorinae
└─o
├─o Troodontidae
└─o Avialae
└─o Aves
Les dinosaures de France
Trias de l'ouest et de l'est de la France : Airel en Manche : squelette de Coelurosaure, Halticosaurus. Pierreclos en Saône-et-Loire : restes de Plateosaurus La Chapoux (anciennement Le Chappou) à Cerin dans le département de l'Ain : Thecodontosaurus. Salins, Arbois, Poligny, Moissey et Domblans dans le Jura : Plateosaurus, Prosauropodes. Violot et Provenchères-sur-Meuse dans la Haute-Marne : Plateosaurus Lunéville et Saint-Nicolas-de-Port dans l'est de la Meurthe-et-Moselle : prosauropodes du genre Plateosaurus et Thecodontosaurus, dents de Coelurosaure.
Airel en Manche : squelette de Coelurosaure, Halticosaurus.
Pierreclos en Saône-et-Loire : restes de Plateosaurus
La Chapoux (anciennement Le Chappou) à Cerin dans le département de l'Ain : Thecodontosaurus.
Salins, Arbois, Poligny, Moissey et Domblans dans le Jura : Plateosaurus, Prosauropodes.
Violot et Provenchères-sur-Meuse dans la Haute-Marne : Plateosaurus
Lunéville et Saint-Nicolas-de-Port dans l'est de la Meurthe-et-Moselle : prosauropodes du genre Plateosaurus et Thecodontosaurus, dents de Coelurosaure.
Jurassique normand, picard ou jurassien Rivages normands, embouchure de la Seine et falaises du Pays de Caux, en particulier au voisinage de Caen, Villers-sur-Mer, Dives, Honfleur, du Havre : dinosaures, gros théropodes, Megalosaurus, Piveteausaurus ou Streptospondylus, coelurosaure, stégosaure Lexovisaurus… Boulogne-sur-Mer et le Boulonnais : restes des sauropopdes, mégalosauridés, coelurosaures, Camptosaurus (ornithopode primitif) Damparis dans le Jura : restes de gros sauropode du genre Bothriospondylus, mégalosauridés…
Rivages normands, embouchure de la Seine et falaises du Pays de Caux, en particulier au voisinage de Caen, Villers-sur-Mer, Dives, Honfleur, du Havre : dinosaures, gros théropodes, Megalosaurus, Piveteausaurus ou Streptospondylus, coelurosaure, stégosaure Lexovisaurus…
Boulogne-sur-Mer et le Boulonnais : restes des sauropopdes, mégalosauridés, coelurosaures, Camptosaurus (ornithopode primitif)
Damparis dans le Jura : restes de gros sauropode du genre Bothriospondylus, mégalosauridés…
Transition Jurassique-Crétacé Canjuers dans le Var : squelette du petit Compsognathus dans les calcaires lithographiques (1971)
Canjuers dans le Var : squelette du petit Compsognathus dans les calcaires lithographiques (1971)
Crétacé inférieur : Angeac en Charente : restes de sauropodes et ornithomimosaures ("dinosaures autruches"). Pays de Bray et Villers-Saint-Barthélemy dans l'Oise : restes de sauropodes Wimereux en Pas-de-Calais : mégalosauridés Wassy en Haute-Marne, Cousancelles, Ville-sur-Saulx en Meuse, Louppy-le-Château, Varenne, Grandpré en Ardennes : mégalosauridés type Erectopus, iguanodons et iguanodontidés… Bedoin et Mondragon dans le Vaucluse : petits sauropodes Fons et Serviers-et-Labaume dans le Gard : dents de thérapode de la famille des deinonychosauridés.
Angeac en Charente : restes de sauropodes et ornithomimosaures ("dinosaures autruches").
Pays de Bray et Villers-Saint-Barthélemy dans l'Oise : restes de sauropodes
Wimereux en Pas-de-Calais : mégalosauridés
Wassy en Haute-Marne, Cousancelles, Ville-sur-Saulx en Meuse, Louppy-le-Château, Varenne, Grandpré en Ardennes : mégalosauridés type Erectopus, iguanodons et iguanodontidés…
Bedoin et Mondragon dans le Vaucluse : petits sauropodes
Fons et Serviers-et-Labaume dans le Gard : dents de thérapode de la famille des deinonychosauridés.
Crétacé supérieur du Languedoc, de la Provence ou de la façade ouest : Saint-Martory dans la Haute-Garonne : fragment de mandibule de hadrosauridés (identifiés après 1975) Villeveyrac, Saint-Chinian, Montpellier et Argelliers dans l'Hérault, Albières et Fa dans l'Aude : iguanodondidés Rhabdoton, hadrosauridés Orthomerus, sauropodes Hypselosaurus et Titanosaurus, ankylosaure Struthiosaurus, petits et grands thérapodes… Campagne-sur-Aude dans l'Aude : Squelette relativement complet d'Ampelosaurus atacis (Titanosauridae) (2000) Aix-en-Provence et contrées voisines dans les Bouches-du-Rhône et le Var : gisements d'œufs de dinosaures Près du Phare de Chassiron, île d'Oléron, Veillon en Vendée : empreintes de dinosaures, spectaculaires sur le dernier site…
Saint-Martory dans la Haute-Garonne : fragment de mandibule de hadrosauridés (identifiés après 1975)
Villeveyrac, Saint-Chinian, Montpellier et Argelliers dans l'Hérault, Albières et Fa dans l'Aude : iguanodondidés Rhabdoton, hadrosauridés Orthomerus, sauropodes Hypselosaurus et Titanosaurus, ankylosaure Struthiosaurus, petits et grands thérapodes…
Campagne-sur-Aude dans l'Aude : Squelette relativement complet d'Ampelosaurus atacis (Titanosauridae) (2000)
Aix-en-Provence et contrées voisines dans les Bouches-du-Rhône et le Var : gisements d'œufs de dinosaures
Près du Phare de Chassiron, île d'Oléron, Veillon en Vendée : empreintes de dinosaures, spectaculaires sur le dernier site…
Terrains tertiaires comportant des sédiments mésozoïques remaniés par la mer des *****s ***** de l'Anjou : dents d'hadrosaures, théropodes.
***** de l'Anjou : dents d'hadrosaures, théropodes.
Musées
De très nombreux musées accueillent des squelettes ou des reconstitutions de dinosaures. Voici une liste non-exhaustive des musées les plus importants pour quelques pays du globe :
Belgique : Muséum des sciences naturelles de Belgique (Bruxelles) Musée de l'Iguanodon de Bernissart
Muséum des sciences naturelles de Belgique (Bruxelles)
Musée de l'Iguanodon de Bernissart
Canada : Royal Tyrrell Museum of Palaeontology (Drumheller, Alberta)
Royal Tyrrell Museum of Palaeontology (Drumheller, Alberta)
États-Unis : American Museum of Natural History (New York) Field Museum (Chicago)
American Museum of Natural History (New York)
Field Museum (Chicago)
France : Galerie d'Anatomie comparée et de Paléontologie - Muséum national d'histoire naturelle Muséum national d'histoire naturelle (Paris) Musée des Confluences (Lyon) Muséum d'histoire naturelle du Var (Toulon) Muséum d'histoire naturelle de Toulouse Muséum d'histoire naturelle d'Aix-en-Provence Muséum d'histoire naturelle du Mans Musée des dinosaures d'Espéraza (Aude) Musée du Parc de la Plaine des Dinosaures de Mèze (Hérault) Musée de l'ACAP de Cruzy (Hérault)
Galerie d'Anatomie comparée et de Paléontologie - Muséum national d'histoire naturelle Muséum national d'histoire naturelle (Paris)
Musée des Confluences (Lyon)
Muséum d'histoire naturelle du Var (Toulon)
Muséum d'histoire naturelle de Toulouse
Muséum d'histoire naturelle d'Aix-en-Provence
Muséum d'histoire naturelle du Mans
Musée des dinosaures d'Espéraza (Aude)
Musée du Parc de la Plaine des Dinosaures de Mèze (Hérault)
Musée de l'ACAP de Cruzy (Hérault)
Royaume-Uni Muséum d'histoire naturelle de Londres (Londres)
Muséum d'histoire naturelle de Londres (Londres)
Suisse Muséum d'histoire naturelle de Genève (Genève)
Muséum d'histoire naturelle de Genève (Genève)
Les dinosaures dans la culture
Les dinosaures dans les arts et la littérature
Gertie le dinosaure est le premier personnage de dessin animé doté d’une personnalité propre, créé en 1904 par le dessinateur Winsor McCay.
Jurassic Park : roman et film mettant en scène des dinosaures, de façon assez réaliste.
Sur la terre des dinosaures (Walking with dinosaurs) : série en six épisodes de Tim Haines, faisant largement appel aux images de synthèse, produite par la BBC en 1999, souvent rediffusée par la chaîne Planète.
Dinosaure est un film de Walt Disney qui mêle créations en images de synthèse et décors réels.
Dinotopia : Livres illustrés pour enfants puis nouvelles pour adolescents, puis romans pour adultes et enfin série télévisée.
Casimir : personnage principal de L'Île aux enfants, une émission de télévision pour la jeunesse dans les années 1970 en France.
Entre mystère et légende, le monstre du Loch Ness fait couler beaucoup d'encre.
Le Monde perdu, un livre de sir Arthur Conan Doyle.
Yoshi : personnage de jeux vidéo de l'univers de Mario.
Denver, le dernier dinosaure, est une série télévisée d'animation franco-américaine mettant en scène un dinosaure né de nos jours.
Le Petit Dinosaure, est une série de dessins animés produits par Steven Spielberg et George Lucas.
Nabuchodinosaure, est le dinosaure improbable et humoristique de la bande-dessinée du même nom.
Ptéra : monstre du jeu vidéo Pokémon.
Rexillius : monstre de pokémon
Kyōryū Sentai Zyuranger : série de sentai dont le premier mot du titre original signifie dinosaure en japonais.
Gon : bébé tyrannosaure, héros d'un manga de Masashi Tanaka.
Nick Cutter et les Portes du temps : série comportant 5 saisons mettant en scène des dinosaures venant de portails spatio-temporel.
Terra Nova : série américaine mettant en scène des individus venant vivre dans le crétacé supérieur.
Les Dinosaures en bande dessinée : bande dessinée visant à faire découvrir le monde des dinosaures sur un ton humoristique.
Future War : film américain de 1997, d'Anthony Doublin.
 词典释义:
词典释义: