 词典释义:
词典释义:
 (
(
 , 无
, 无 源说)
源说) 词
词 物学, 植物
物学, 植物 物学;
物学; 作用,光
作用,光

 ;
; 育;
育; 物化学;
物化学; 物学】无
物学】无
 源说, 自然
源说, 自然
 说
说
 源[说、论];
源[说、论];  物自
物自 论; 非
论; 非 物
物 因
因L'abiogenèse, dans son sens le plus courant, désigne l'étude de la génération de la vie à partir de la matière non vivante. Aujourd'hui, le terme est utilisé principalement en biologie, dans le contexte de l'origine de la vie.
Les origines de la vie sur Terre demeurent incertaines. Il y a de nombreuses théories scientifiques qui tentent d'expliquer l'apparition de la vie qui remonte au précambrien. Une étude publiée en 2015 envisage qu'une forme embryonnaire de vie pourrait être apparue il y a près de 4,1 milliards d'années.
Stromatolites du précambrien, dans la formation de Siyeh. En 2002, William Schopf a affirmé que ces formations étaient âgées de 3,5 milliards d'années, elles seraient alors les plus anciennes traces de vie sur Terre.
Stromatolites dans la Baie Shark (Australie).
Étapes vers l'apparition de la vie
Il n'existe pas de modèle « standard » pour décrire l'origine de la vie. En fait il y a actuellement deux écoles de pensée principales sur l'origine de la vie :
l’une (fondée sur la théorie de l'information et les problématiques de codage de l'information) qui considère que seuls les acides nucléiques ont pu jouer un rôle clé dans le processus de la sélection naturelle qui a formaté les organismes vivants ;
l’autre qui privilégie le rôle du métabolisme, qui considère que les organismes vivants sont essentiellement des systèmes loin de leur état d’équilibre thermodynamique, qui parviennent à se maintenir tels grâce aux processus métaboliques.
Les modèles « gènes d'abord »
Dans ce modèle, l'apparition du génome a précédé l'apparition du métabolisme. Des molécules d'ADN ou d'ARN auraient ainsi existé seules, s'autorépliquant à partir des molécules présentes dans leur environnement. Les « individus » sont donc représentés par les molécules d'acides nucléiques elles-mêmes.
L’école de pensée privilégiant les acides nucléiques se subdivise en deux classes d’hypothèses :
la première est que les premières formes de vie sur Terre ont utilisé l’ARN comme seul constituant génétique capable d’encoder l’information nécessaire aux catalyses biologiques ;
la deuxième est encore plus restrictive puisqu’elle considère l’ARN comme la première forme de vie sur Terre, c’est-à-dire comme le premier système chimique à l’origine de l’évolution darwinienne, l'hypothèse du monde à ARN.
Les modèles « métabolisme d'abord »
Plusieurs modèles rejettent l'idée de l'autoréplication d'un gène « nu » et font l'hypothèse de l'apparition d'un métabolisme primitif qui aurait précédé l'émergence de la réplication de l'ARN. Une des premières versions de cette hypothèse fut présentée en 1924 par Alexander Oparin avec son idée de vésicules primitives capables de se répliquer (coacervats), à une époque où on ne connaissait pas encore la structure de l'ADN.
L’école de pensée privilégiant le rôle du métabolisme, comme le concept d’autopoïèse, s’inspire du concept du système dissipatif qui se maintient loin de l'équilibre thermodynamique, grâce au flux de matière et d'énergie qui le traversent.
Le problème de cette approche est de pouvoir rendre compte de l'évolution de ces systèmes métaboliques, car ils se répliquent sans transmettre d’information, c’est-à-dire qu’ils n'ont pas d'hérédité.
D'autres variantes sont apparues dans les années 1980 et 1990 comme la théorie de Günter Wächtershäuser sur un monde sulfuro-ferreux, ou les modèles de Christian de Duve fondés sur la chimie des thioesters.
D'autres arguments plus abstraits ont aussi été présentés. On peut citer les modèles mathématiques de Freeman Dyson au début des années 1980 sur la probabilité de l'émergence d'un métabolisme sans présence de gènes, ou encore les travaux de Stuart Kauffman sur les ensembles globalement autocatalytiques (voir génération spontanée pour une présentation des idées de Kauffman sur l'origine de la vie).
Principales étapes
Le modèle le plus couramment accepté est fondé sur l'enchaînement supposé des événements suivants :
Formation de la « soupe primitive ». Des conditions prébiotiques plausibles entraînent la création de molécules organiques simples, qui sont les briques de base du vivant. Les réactions de synthèses organiques conduisant à la formation de catalyseurs favorables à ces mêmes synthèses bénéficient d'un avantage sélectif. Suivant les auteurs, des mécanismes différents expliquent la formation des monomères initiaux, et leur polymérisation ultérieure.
Cloisonnement et compartimentation des milieux réactifs. Des phospholipides forment spontanément des doubles couches qui sont la structure de base des membranes cellulaires. L'organisation spatiale permet des différences de compositions entre milieux, et la mise en place d'échanges trans-membranaires. Apparition de cycles réactifs à la base d'un métabolisme élémentaire, capable d'alimenter un processus de production et reproduction du milieu chimique en puisant sur des ressources extérieures ; avantage sélectif aux collections de cycles capables de produire leur propres catalyseurs.
Réplication de molécules à la fois fonctionnelles et codantes. Les mécanismes qui produisent aléatoirement des molécules d'ARN (acide ribonucléique), en mesure d'agir comme des ARN-enzymes capables, dans certaines conditions très particulières, de se dupliquer. C'est une première forme de génome, et nous sommes alors en présence de protocellules. Grâce à une transcription codée des éléments participant au métabolisme d'ensemble, les compartiments élémentaires sont capables de maintenir une composition chimique et un métabolisme stable, de dupliquer les molécules codantes qui régulent ce milieu, autorisant la séparation d'une cellule initiale en deux cellules fonctionnellement équivalentes.
Spécialisation de la fonction enzymatique et de la réplication. Les ARN-enzymes sont progressivement remplacées par des protéines-enzymes, grâce à l'apparition des ribozymes, ceux-ci étant capables de réaliser la biosynthèse des protéines. L'ADN apparaît et remplace l'ARN dans le rôle de support du génome, dans le même temps les ribozymes sont complétés par des protéines, formant les ribosomes. C'est l'apparition de l'organisation actuelle des organismes vivants.
Les ARN-enzymes sont progressivement remplacées par des protéines-enzymes, grâce à l'apparition des ribozymes, ceux-ci étant capables de réaliser la biosynthèse des protéines.
L'ADN apparaît et remplace l'ARN dans le rôle de support du génome, dans le même temps les ribozymes sont complétés par des protéines, formant les ribosomes. C'est l'apparition de l'organisation actuelle des organismes vivants.
Cette logique d'apparition correspond au modèle « métabolisme d'abord ». Les étapes 2 et 3 sont parfois inversées, l'isolement en compartiment et l'émergence du métabolisme étant alors présenté après l'apparition des ARN autoréplicants (hypothèse « gènes d'abord »).
Kauffman et les réseaux autocatalytiques
Dans le Jeu de la vie des formes stables et répétitives peuvent apparaître, et tendent à être privilégiées dans la durée par rapport aux formes évanescentes.
Que des formes complexes ne puissent pas avoir une origine aléatoire était un principe très généralement accepté. En 2002, Stuart Kauffman est le premier à l’avoir mis en doute à la suite d’expériences numériques, point de vue qu'il défendit dans l'introduction de son ouvrage Self Organization in Biological Systems. Il étudiait la dynamique de réseaux d’automates cellulaires engendrés d’une façon aléatoire. Il a alors observé que des formes stables animées d’un mouvement périodique apparaissaient spontanément, quelles que soient les conditions initiales. Il voyait ainsi des formes complexes et durables engendrées d’une façon purement aléatoire. Il a vite compris que son observation renouvelle d’une façon très originale nos façons d’expliquer les phénomènes et qu’il pouvait l’appliquer à la question de l’origine de la vie. Il a mis quelques années avant de le faire accepter par d’autres scientifiques.
Lorsque de nombreuses molécules différentes sont mises en présence, on peut décrire la dynamique chimique par un réseau. Chaque espèce de molécule est reliée aux autres espèces avec lesquelles elle réagit. Un tel réseau peut avoir des boucles autocatalytiques, c’est-à-dire qu’une espèce moléculaire, appelons-la A, favorise, ou catalyse, des réactions chimiques qui vont conduire à la production de molécules du type A. Une telle boucle est potentiellement explosive, parce qu’il y a un effet boule de neige, mais si les espèces à partir desquelles A est produite sont en quantité limitée, l’autoproduction de A est elle-même limitée.
Les êtres vivants sont toujours des réseaux autocatalytiques. Toutes leurs molécules (ADN, protéines et autres) réagissent avec les molécules ingérées (les aliments) pour produire des molécules semblables à elles-mêmes. Les plantes sont même capables d’utiliser la lumière du soleil dans ce but.
En étudiant des modèles, Kauffman a établi que sous des conditions assez générales un réseau de réactions chimiques contient nécessairement des réseaux autocatalytiques. Il suffit que le réseau soit suffisamment touffu, que les espèces moléculaires soient suffisamment nombreuses et réactives les unes vis-à-vis des autres.
Les réseaux autocatalytiques expliquent la reproduction des molécules mais à eux seuls ils ne suffisent pas pour expliquer la reproduction des unicellulaires.
Chimie prébiotique
La chimie prébiotique est une branche de la biochimie qui se propose d'étudier les conditions primitives de la vie sur Terre, et surtout la manière dont elle est apparue à partir de quelques molécules inertes.
Conditions chimiques initiales
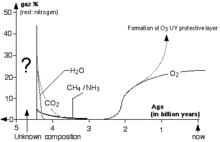
L’atmosphère de l’Archéen ne contient apparemment pas ou très peu d’oxygène libre. Selon Alexandre Oparine et John Haldane, l'atmosphère terrestre primitive, lors de sa formation, était composée de méthane (CH4), d'ammoniac (NH3), de vapeur d'eau (H2O), de gaz carbonique (CO2) et d'hydrogène sulfuré (H2S). Par l'action du rayonnement ultraviolet provoquant la photodissociation de ces molécules, l'atmosphère a évolué progressivement, perdant son méthane pour évoluer vers une atmosphère de gaz carbonique et d'azote. Lors de l'apparition de la vie, l'atmosphère est moyennement réductrice : (CO2 ; N2). Des modèles plus anciens considéraient une atmosphère fortement réductrice comme probable : (CH4 ; NH3).
Sa température est supérieure à celle d’aujourd’hui, bien que le Soleil soit de 25 à 30 % moins lumineux que de nos jours, la différence est compensée par la présence de gaz à effet de serre, ou, alternativement, par l'absence de nuages réfléchissants et un albédo de surface plus bas. La pression est de quelques atmosphères.
La température dans la majorité des modèles est de 40 °C à 85 °C, bien qu’une atmosphère plus tempérée soit possible. L’eau sous forme liquide est présente, les océans ont probablement fini de se former durant l’Hadéen.
L'expérience Urey-Miller et l'origine des molécules organiques
Le biochimiste Robert Shapiro a résumé les théories de Oparin et Haldane sur la « soupe primordiale » de la manière suivante :
La Terre avait primitivement une atmosphère réductrice.
Cette atmosphère a produit des composés organiques simples (monomères) sous l'action de diverses sources d'énergie.
Ces composés se sont accumulés au fil du temps dans une sorte de soupe chimique, qui a pu se trouver concentrée en divers endroits (côte et lagune, mont hydrothermal, etc.).
Par la suite, des polymères et composés plus complexes se sont développés dans le mélange, conduisant finalement à la Vie.
Les trois premières étapes ont fait l'objet de reconstitutions et d'études en laboratoire ; mais la quatrième « étape » de cette théorie a été qualifiée de très simpliste - rappelant le coup de baguette magique des contes de fées.
En 1953, le chimiste américain Stanley Miller réalisa une série d'expériences dans le cadre de sa thèse de doctorat.
Les expériences de Miller-Urey furent décisives dans la compréhension de la chimie prébiotique. En effet, à partir d'eau (H2O), de méthane (CH4), d'ammoniac (NH3) et d'hydrogène (H2), on a recueilli des acides aminés (dont 13 des 22 qui sont utilisés pour fabriquer des protéines dans les cellules des organismes), des sucres, des lipides, et quelques composants des acides nucléiques mais pas d'acides nucléiques entiers (ADN ou ARN). Des racémiques des énantiomères gauche lévogyre et droite dextrogyre se sont formés.
Les travaux révolutionnaires de Miller ont donné lieu à de très nombreuses publications, la composition de l'atmosphère terrestre primitive faisant débat.
En 2009, Sutherland et son équipe sont parvenus à établir une voie de synthèse des nucléotides à base pyrimidique, l'acide uridylique et l'acide cytidylique. Les chercheurs butaient sur cette voie de synthèse prébiotique depuis 40 ans en raison de la difficulté à trouver la bonne façon de lier le ribose à la base azotée. La clé de cette voie est de passer par un précurseur commun au ribose et la base azotée. Ce précurseur, le 2-aminooxazol, est obtenu à partir de molécules organiques élémentaires : le glycéraldéhyde, le cyanamide, le cyanoacétaldéhyde, le cyanoacétylène et le phosphate inorganique. Le mélange réactionnel alimenté par de l'azote gazeux est soumis à un cycle de chauffage-refroidissement afin de simuler le cycle d'évaporation d'une mare par le rayonnement solaire et l'alimentation par la pluie. Après une semaine le 2-aminooxazol s'accumule dans le réacteur. Le précurseur se transforme ensuite en ribose et cytosine liés ensemble. Le phosphate est ensuite ajouté au milieu réactionnel en présence d'UV durant trois jours (l'absence d'ozone dans les conditions prébiotiques engendrait un bombardement intense d'UV). L'acide cytidylique se synthétise sous l'effet du rayonnement UV et quelques nucléotides portant une cytosine se transforment en acide uridylique. Pour les deux autres nucléotides, l'équipe de Sutherland travaillait sur un précurseur commun aux acides nucléiques à base purique.
La formation de molécules organiques se rencontre également dans l'Atmosphère de Titan.
Asymétrie des biomolécules
La chimie prébiotique conduit a priori à un racémique de molécules. Or, on sait depuis le milieu du XIX siècle (notamment avec des travaux de Pasteur en 1847) que les acides aminés et les glucides simples naturels n'existent pratiquement que sous une de leurs deux formes énantiomères : la forme L pour les premiers et la forme D pour les seconds. On parle d'homochiralité du vivant. Si l'on suppose que cette asymétrie dans les composés monomères remonte à la formation de la « soupe originelle », se pose alors la question de l'origine de cette énantiospécificité originelle.
Cette asymétrie des molécules biologiques a été utilisée comme argument en faveur d'un dessein intelligent de l'origine de la vie.
Si l'on admet en revanche que l'énantiospécificité n'intervient qu'avec la formation de polymères, elle peut recevoir une explication plus simple. Les polypeptides sont plus stables quand ils sont formés par des acides aminés de même forme, qu'ils soient tous L ou tous D. Une chaîne composée d'un mélange des deux sortes d'isomères ne serait pas stable, parce que quand un acide aminé L est associé à un D, les deux chaînes latérales dépassent du même côté, conduisant à une tension structurelle qui affaiblit la liaison chimique.
Du moment que l'avantage sélectif appartient aux cycles qui produisent leurs propres catalyseurs, il suffit que le premier polypeptide impliqué dans un cycle favorable ait été composé de L pour que sa synthèse elle-même ait par la suite favorisé ce type de composé.
Polymérisation : condensation sur surfaces minérales

Le scénario de la « soupe primitive » comportant des micelles ayant des limites, les scientifiques commencèrent à repenser l'évolution de la vie prébiotique sans membrane, et imaginèrent des molécules organiques capables de croître sur une surface minérale. Les chimistes Graham Cairns-Smith et Günter Wächtershäuser (en), montrèrent que les argiles et les cristaux de pyrite permettent l'agglutination des molécules organiques facilement. Une surface est plus stable qu'un espace tridimensionnel comme l'eau, mais aussi plus réduit (deux dimensions). Dans un espace tridimensionnel, les molécules sont sujettes à des contraintes selon tous les axes. Ainsi, à la surface des cristaux le milieu devient très réactif et facilite l'agglomération des molécules organiques.
L'assemblage de petites molécules (comme les acides aminés) en macromolécules (comme les protéines) nécessite l'élimination de molécules d'eau. Or, la thermodynamique indique qu'il est défavorable de réaliser une telle condensation dans l'eau elle-même. Il est possible pour résoudre cette contradiction de faire appel à des surfaces minérales, comme les micas, les argiles ou les pyrites. L'adsorption des petites molécules sur ces surfaces les concentre et les modifie chimiquement, ce qui peut rendre la formation de macromolécules plus favorable.
L'argile, par exemple, se trouve très abondamment sur Terre et est constituée d'un empilement de couches fines. Entre les différentes couches de l'argile peuvent se glisser certaines petites molécules organiques, ce qui permet une adsorption importante. L'argile est aussi un catalyseur très efficace pour de nombreuses réactions organiques, et aurait donc pu permettre la polymérisation des acides aminés et/ou des acides nucléiques. Le chimiste anglais Graham Cairns-Smith a développé cette hypothèse dans Seven clues to the origin of life en 1985 (traduction française : L'énigme de la vie, 1990).
Le prix Nobel de chimie Thomas Cech indiqua que l'ARN pouvait être la première molécule vivante du fait de ses propriétés catalytiques et auto-catalytiques (on pensait que seules les protéines-enzymes en étaient capables).
Pression de sélection sur les molécules complexes
La « soupe primordiale » est initialement formée par des molécules relativement simples, mais sa composition se complexifie progressivement suite aux réactions chimiques portant sur les molécules de première génération. D'une part, l'action de la source d'énergie (rayonnement ultraviolet ou autre) sur ces premières molécules formées créé des radicaux libres plus complexes, capables d'interagir avec les molécules de premières générations pour conduire par génération successives à des molécules de plus en plus lourdes. D'autre part, les molécules ainsi formées peuvent directement réagir entre elles, conduisant à de nouvelles synthèses.
Dans un premier temps, l'évolution de la composition ne dépend que de la constante d'équilibre et de la concentration molaire des paires de molécules susceptibles d'interagir. Cependant, dans un deuxième temps, certaines des molécules formées peuvent intervenir comme catalyseur sur telle ou telle réaction. À partir du moment où l'effet d'un catalyseur porte sur des réactions chimiques qui conduisent (directement ou non) à sa propre formation, une réaction en chaîne est susceptible de se mettre en place. Cette rétroaction chimique déplace l'équilibre global de la composition, augmentant progressivement la concentration du catalyseur, et tendant à assécher les réactifs consommés, qui ne peuvent alors plus alimenter d'autres types de synthèses.
Ainsi, avant même qu'un code génétique puisse se mettre en place, une pression de sélection a pu s'exercer sur la formation des molécules plus complexes, ou sur la formation de groupe fonctionnel au sein de ces molécules. Les réactions de synthèses organiques conduisant à la formation de catalyseurs favorables à ces mêmes synthèses bénéficient d'un avantage sélectif.
À un premier niveau, cette pression de sélection chimique ne fait initialement que favoriser la formation de molécules et de groupes fonctionnels résultant de voies de synthèse relativement simples.
Cependant, des chaînes polymérisées formées de monomères de même famille ont pu être synthétisées dans ce milieu. Dans des conditions chimiques favorables, certaines de ces chaînes polymérisées peuvent servir de modèles pour leur propre réplication, devenant ainsi leur propre catalyseur. Si l'on suppose que le milieu chimique est favorable et contient les constituants nécessaires, non seulement à la synthèse de ces polymères, mais également à leur réplication, les réactions chimiques de premier niveau, présentes dans la « soupe primordiale » initiale, conduisent alors, à un second niveau, à alimenter non seulement la formation, mais également la réplications de chaînes polymérisées.
Ces chaînes évoluent par mutation, peuvent se complexifier, et certaines des chaînes formées peuvent à leur tour intervenir comme catalyseur sur telle ou telle réaction de premier niveau. Ceci conduit à un second niveau de rétroaction, où des collections de chaînes polymérisées servent de catalyseurs aux différentes réactions nécessaires pour la synthèse et l'assemblage de leurs constituants. Cette « pré-vie » est capable de sélection et de mutation, mais n'est pas encore capable à ce niveau de stabiliser ni un métabolisme défini, ni une réplication fidèle.
Métabolisme et génome : un monde à ARN
S'il est logique de penser que des polymères non homogènes aient pu faire l'objet d'une pression de sélection, la nature exacte de ces polymères est une question ouverte quand on étudie la biochimie actuelle. On a pu penser que les polymères primitifs ont pu être les peptides (polymères d'acide aminé), privilégiant le rôle primordial des polypeptides dans le métabolisme. On a pu au contraire penser que les polymères primitifs étaient ceux de l'ADN, privilégiant sa fonction de réplication. Mais ces deux aspects sont inter-dépendants, et mutuellement exclusifs ; alors que la logique d'une approche de type abiogenèse suppose l'existence d'un polymère à la fois auto-répliquant et ayant des fonctions de catalyseur.
L'hypothèse du monde à acide ribonucléique (ARN) est que l'ARN était la principale — et sans doute la seule — forme de vie avant l'émergence de la première cellule à ADN. C'est Walter Gilbert qui a utilisé pour la première fois le terme « monde à ARN » (en anglais : RNA world) en 1986.
En 1990, Larry Gold et Jack Szostak ont mis au point une méthode visant à diriger l'évolution d'ARN, afin de sélectionner ceux montrant une activité catalytique. Ils ont depuis réussi à obtenir des ribozymes capables de lier des nucléotides entre eux, de lier des acides aminés à des ARN, d'effectuer des réactions d'oxydo-réductions, de se lier à des composants de la membrane, etc.
Il est donc en théorie possible, sur ce modèle, que l'ARN seul suffise à établir un métabolisme primitif. Toutefois, il reste encore à découvrir un ARN capable de se répliquer lui-même.
L'hypothèse d'un monde à ARN a aujourd'hui la faveur des scientifiques et est fondée sur plusieurs éléments. Notamment sur le fait que l'ARN est en théorie capable aussi bien d'assurer des tâches métaboliques que d'être le support d'une information génétique.
Des molécules organiques aux protocellules
Formation spontanée des vésicules
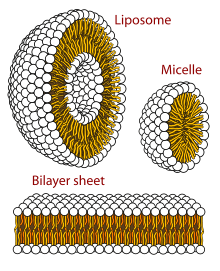
Il suffit d’agiter une eau savonneuse pour obtenir de la mousse. L’écume de la mer se forme de la même façon, d’autant plus facilement que l’eau est plus sale. On l’explique à partir du caractère amphiphile de certaines molécules. Une molécule est hydrophile si elle « préfère » être dans l’eau que dans l’huile, c’est-à-dire si spontanément, du fait des lois de la physique statistique, elles se concentrent davantage dans l’eau que dans l’huile. Elle est hydrophobe, ou lipophile, dans le cas inverse. Certaines molécules, telles que celles du savon, sont allongées avec une extrémité hydrophile et l’autre hydrophobe, c’est pourquoi elles sont dites amphiphiles. Lorsqu’elles sont mises en solution dans l’eau, elles forment alors spontanément de nombreuses structures qui peuvent être très complexes et notamment des bicouches qui se replient en vésicules. Une bicouche est une membrane, plongée dans l'eau, dont les deux faces sont composées de têtes hydrophiles, les extrémités hydrophobes étant rassemblées à l’intérieur de la bicouche. Une vésicule est un petit sac plein d’eau, qui flotte dans l’eau, et dont la membrane est une bicouche.
Les cellules des êtres vivants sont des vésicules très élaborées. Leur membrane est essentiellement une bicouche lipidique, mais elle est beaucoup plus complexe que les membranes des vésicules dans l’eau savonneuse. L’intérieur de la cellule surtout est très différent de son extérieur, ce qui n’est pas le cas des vésicules que l’on forme en agitant une eau sale.
Importance de la frontière dans la définition du soi
Schéma d'une cellule animale. (1) Nucléole (2) Noyau (3) Ribosomes (4) Vésicule (5) Réticulum endoplasmique rugueux (ou granuleux) (REG) (6) Appareil de Golgi (7) Cytosquelette (8) Réticulum endoplasmique lisse (9) Mitochondries (10) Vacuole (11) Cytosol (12) Lysosome (13) Centrosome (constitué de deux centrioles) (14) Membrane plasmique
Aujourd'hui, de nombreux modèles résolvent le problème de l'apparition des molécules organiques. Les scientifiques arrivent à produire de nombreuses petites molécules biologiques (acides aminés, sucres, bases nucléiques) dans des conditions prébiotiques en laboratoires. Cependant, les expériences de Miller et les modèles qui en sont dérivés ne fournissent pas d'explication sur les étapes suivantes qui incluent la transition des monomères aux biopolymères, puis aux protocellules et finalement aux cellules vivantes ayant un métabolisme de base. Aussi les scientifiques ont exploré d'autres voies de recherche.
Dans l'enquête sur l'origine de la vie, on cherche ce qui a pu donner à « une entité » la capacité de « se » reproduire. Une telle capacité suppose logiquement, en amont, d'avoir réalisé une limite entre ce qui est « soi » et le reste du monde. Cette limite intervient dans l'espace, sous la forme d'une frontière ; elle intervient également dans la nature de ce qui est reproduit, associé à un certain type de fonctionnement - essentiellement, un métabolisme. La racine essentielle d'un être est sa capacité à maintenir en permanence la cohérence de ses caractéristiques (ce qui constitue sa définition) dans le temps et dans l'espace. Cette permanence étant réalisée par l'entité elle-même, on parle d’autopoïèse pour caractériser une telle situation (du grec auto soi-même, et poièsis production, création).
L’exemple canonique d’un système autopoïétique fourni par Francisco Varela et Humberto Maturana est celui de la cellule biologique. Le modèle minimal d'un système autopoïétique est une cellule dont la membrane est composé d'un constituant C se dégradant en D et baignant dans un milieu riche en molécules A. Ces molécules A peuvent franchir la membrane et être transformées par le métabolisme régnant au sein de la cellule en molécules B, pour lesquelles la membrane est imperméable. B peut s'intégrer à la membrane pour se transformer en C. Si le débit d'entrée de A et sa conversion en B sont suffisamment grands devant le coefficient de dégradation de C en D, alors la cellule se maintient au cours du temps.
Ce modèle montre l'importance de la frontière du système (ici la membrane) : si celle-ci disparaît, le métabolisme et le système entier s'effondre. Il s'agit d'un cercle vicieux : si B s'échappe, sa concentration diminue de telle sorte que la membrane se dégrade de plus en plus vite et que la perte en B augmente. Le métabolisme et la membrane dépendent l'un de l'autre, la structure ne peut se maintenir sans le flux.
Structures cellulaires en émulsion
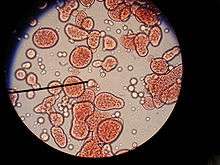
Les expériences sur la formation spontanée de membranes cellulaires sont bien antérieures aux synthèses de Urey-Miller de 1953.
Après de multiples réflexions et hypothèses, le biochimiste soviétique Alexandre Oparine est parvenu à créer de petites structures sphériques appelées coacervats à partir d'une émulsion contenant des protéines histones et de la gomme arabique, en 1922. Il s'agissait de sphérules colloïdales, de quelques micromètres de diamètre en suspension et liées entre elles. Les savons et les émulsions sont des substances de type colloïde.
Les coacervats d'Oparine ont la propriété d'être séparés du monde par une surface hydrophobe polyhémiacétalique (ils ont un intérieur et un extérieur) tout comme les cellules biologiques (mais dans le cas étudié, il ne s'agit pas de lipides). La similarité ne s'arrête pas là : en présence de catalyseurs, ces coacervats échangeaient des substances avec le monde extérieur, grossissaient et finissaient par se multiplier par scission.
C'est à partir de cette expérience que l'on imagina la synthèse possible d'un système vivant à partir de molécules organiques, d'eau et d'énergie provenant d'un rayonnement électromagnétique (Soleil, éclairs) : la « soupe prébiotique ».
Reprenant ces travaux, l'anglais John Haldane proposa, en 1929, que ces systèmes « biologiques » ne pouvaient évoluer dans une atmosphère oxydante, en présence d'oxygène. En effet, la survie des molécules organiques (glucides, lipides, protéines, etc.) n'est possible qu'en l'absence de leur oxydation par le dioxygène.
Vésicules lipidiques
L'apparition de vésicules dont la membrane serait formée de phospholipides pose de sérieux problèmes car ces lipides n'étaient pas présents sur la Terre primitive. Cependant des molécules amphiphiles plus simples ont pu être synthétisées de manière abiotique dans des conditions particulières. Par exemple il est reconnu que les sources hydrothermales ('mont hydrothermal') sont des environnements favorables pour des synthèses abiotiques soutenues. À des températures entre 100 °C et 400 °C, des composés amphiphiles peuvent être produits par des réactions chimiques semblables au procédé Fischer-Tropsch. De fait, la production abiotique de chaines courtes d'hydrocarbures a été récemment découverte au niveau du site hydrothermal Lost City situé au milieu de l'Atlantique. Les concentrations millimolaires de méthane pourraient être à l'origine des réactions chimiques réductrices du carbone au niveau de ce type de site hydrothermal.
De nombreuses expériences ont montré que les vésicules dont la membrane est formée de lipides amphiphile sont capables de croître et de se diviser à partir de l'insertion additionnelle de nouveaux lipides. L'idée principale de cette théorie est que la composition moléculaire des membranes vésiculaires serait un moyen primitif de stockage d'information, et d'évolution possible vers l'apparition de biopolymères.
Les études réalisées sur des vésicules lipidiques se sont limitées jusqu'à maintenant à des membranes formées de un à deux types de composés amphiphile. Ces conditions expérimentales ne reflètent pas les résultats de simulations de réactions de chimie prébiotique qui, typiquement, produisent un mélange très hétérogène de composés. Dans le cadre de cette hypothèse d'une membrane formée d'une bicouche lipidique composée d'un mélange hétérogène de molécules amphiphile il existe un nombre considérable de combinaisons possibles pour le positionnement respectif des différents types de molécules au sein de la membrane.
Parmi toutes ces combinaisons potentielles il se peut qu'une combinaison, correspondant à un positionnement local spécifique de la partie interne de la membrane, favorise l'émergence d'un hypercycle selon la terminologie de Manfred Eigen. Il s'agit en fait d'une réaction autocatalytique mutuelle entre une molécule, dont la synthèse est catalysée par le positionnement local spécifique, et le site réactif membranaire qui apparaît suite à la stabilisation de la conformation spatiale du positionnement local par la molécule.
À partir d'expériences réalisées sur des vésicules à membrane hétérogène on peut en déduire que les paires site réactif/molécule seraient transmissibles aux vésicules filles. Une telle hérédité pourrait être à l'origine de lignées distinctes de vésicules permettant à la sélection naturelle d'opérer.
Protocellules
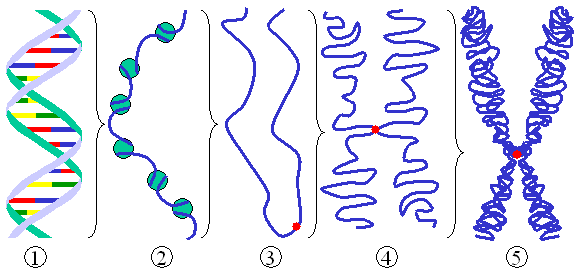
Schéma général d'un chromosome eucaryote.
|
Schéma d'une membrane semi-perméable.
|
De très nombreuses vésicules ont pu se former dans l’océan primitif, aussi facilement qu’aujourd’hui l’écume de la mer. Qu’une telle vésicule soit le lieu de réactions autocatalytiques est tout à fait plausible. Si ses membranes sont telles qu’elles laissent pénétrer les petites molécules nécessaires à la reproduction des grosses, alors on obtient, par le simple jeu des lois physiques et chimiques, un organisme capable de s’alimenter et de grandir. Bien sûr les vésicules ne sont pas toujours dotées d’une telle capacité, mais si elles sont assez nombreuses et assez diversifiées un tel événement n’est peut-être pas complètement improbable.
Un compartiment isolé par une membrane ne forme cependant pas une protocellule. Pour qu’une cellule soit vivante, il ne suffit pas qu’elle soit capable de grandir, il faut encore qu’elle puisse se reproduire. La destinée la plus probable d’une vésicule en croissance est d’exploser et donc de disperser tout son contenu, ce qui revient à mourir. Mais on peut supposer que certaines vésicules aient incorporé à leur paroi des molécules qui la rendent susceptible de former de nouvelles vésicules, par l'intermédiaire de protubérances qui se détachent par exemple. Cette partie du scénario ici proposé de l’origine de la vie n’est peut-être pas très plausible mais elle n’est pas non plus complètement exclue, surtout si l’on songe à la diversité des structures bidimensionnelles qui peuvent se former spontanément sur des bicouches.
Si une vésicule est capable de croître en s’alimentant et de se reproduire, en formant des protubérances, alors elle est un être vivant primitif. Elle a la propriété essentielle des êtres vivants, la capacité de se reproduire quand elle est placée dans un environnement approprié. On peut alors supposer qu’une telle vésicule est l’ancêtre de tous les êtres vivants qui existent aujourd’hui.
Selon Maynard Smith, deux conditions sont nécessaires pour former une véritable protocellule :
Les molécules capables de répliquer la forme de base (les réplicateurs) doivent se lier entre elles en un « chromosome », formant ainsi une unité structurelle, garantissant aux réplicateurs de former un tout cohérent après la réplication ;
la membrane doit posséder des mécanismes d'échange avec le milieu extérieur, autres que les systèmes à protéines actuels.
Selon Wilhelm Reich, des vésicules appelées bions sont apparues lors de ses expériences, à partir de matières minérales et organiques portées à ébullition. Ces vésicules se seraient organisées et auraient donné naissance à des cellules présentant toutes les caractéristiques de la vie. La validité de ces expériences est rejetée par l'ensemble de la communauté scientifique ; les disciples de Reich estiment que ce rejet est le signe d'un complot universel contre leur maître à penser.
Origine de l’ADN et du code génétique
Les premiers réseaux autocatalytiques n’étaient pas aussi élaborés que ceux d’aujourd’hui, fondés sur la machinerie très complexe de l’ADN, des ribosomes, du code génétique et des protéines. Mais les êtres vivants primitifs étaient capables d’évoluer. Leurs réseaux autocatalytiques pouvaient être modifiés par l’incorporation de nouvelles molécules, absorbées de façon exceptionnelle.
De telles modifications sont héritables, parce qu’une fois qu’une molécule est incorporée à un réseau autocatalytique, elle devient capable de se reproduire. Les conditions de l’évolution par la sélection naturelle, telles qu’elles ont été énoncées par Darwin (L'Origine des espèces), sont donc réunies : variations aléatoires héritables et compétition au sein d’une population pour l’accès aux ressources. Les cellules primitives les plus performantes sont celles qui se reproduisaient le mieux et elles tendaient à dominer la population. On peut alors supposer que les êtres vivants primitifs ont évolué. Leurs techniques autocatalytiques rudimentaires se sont perfectionnées jusqu’à atteindre un point de quasi-perfection, à savoir les techniques de réplication de l'ADN et de fabrication des protéines qui sont possédées depuis des milliards d’années par tous les êtres vivants.
De l'ARN à l'ADN
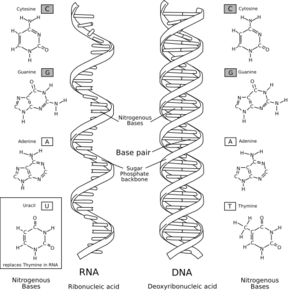
Dans l'hypothèse du monde à ARN, apparaissent tout d'abord des viroïdes ressemblant à des ARN auto-catalytiques, présents dans des compartiments isolés (qu'ils soient membranaires ou cristallins). Puis des protocellules, capables de métabolisme archaïque, sont soumises à une évolution darwinienne, évoluant ainsi vers des cellules à ARN, capables de présenter une activité variée et complexe.
Ribozymes ou protéines ?
Bien que l'ARN soit donc à l'origine de l'ADN dans le métabolisme cellulaire, cette réaction est très difficile à réaliser. De fait, dans les trois lignées, elle est catalysée par des protéines spécialisées : les ribonucléotides-réductases. De plus, cette réaction est très coûteuse en énergie, du fait de la réduction du ribose, et elle produit des radicaux libres, très réactifs, sur la protéine. L'ARN étant une molécule fragile, il paraît improbable qu'elle puisse supporter des radicaux libres sans l'intervention de protéines.
Ainsi, l'origine de l'ADN trouve vraisemblablement sa source après l'apparition des protéines, indispensables à chaque étape de sa synthèse à partir de précurseurs de type ARN, au sein de la cellule.
Intérêt de l'ADN
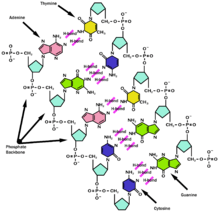
L'ADN présente un certain nombre d'avantages sur l'ARN, en termes de conservation de l'intégrité de l'information génétique.
Tout d'abord il se casse moins facilement, car le désoxyribose de l'ADN contient un atome d'oxygène de moins que le ribose de l'ARN. Or l'oxygène peut facilement interagir sur les liaisons entre nucléotides, posant alors un problème de stabilité.
Certains auteurs avancent l'hypothèse que la désamination spontanée de la cytosine en uracile, aisément détectable dans l'ADN (où U n'est pas normalement présent) par la machinerie cellulaire de réparation des mutations, expliquerait l'utilisation de la base T dans l'ADN. L'ARN (notamment l'ARNm), molécule régulièrement renouvelée dans la cellule, ne voit pas sa séquence contrôlée par des systèmes de réparation, d'où une conservation de la base U dans cette molécule. Dans cette hypothèse, la base U est donc ancestrale, la base T dérivée.
Avantage sélectif de l'ADN : l'hypothèse du virus
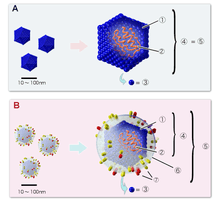
Les avantages en termes de stabilité de l'ADN pourraient ne pas suffire à expliquer son adoption. Ainsi, Patrick Forterre avance l'hypothèse qu'un avantage sélectif supplémentaire peut être dû aux conflits entre virus et cellules vivantes.
Dans ce modèle, le premier organisme à ADN serait un virus. L'ADN conférerait au virus le pouvoir de résister à des enzymes dégradant les génomes à ARN, arme de défense probable des cellules. On retrouve le même principe chez des virus actuels, qui altèrent leur ADN pour résister à des enzymes produites par des bactéries infectées.
Actuellement, on peut observer que les enzymes nécessaires à la traduction de l'ARN vers l'ADN sont très présentes chez les rétrovirus, dont le génome est porté par de l'ARN. De la même façon, de nombreux virus codent leurs propres enzymes de synthèse de l'ADN.
Cette hypothèse est également corroborée par la découverte de virus à ADN, dont celui-ci contient, non pas des groupements thymines, mais des groupements uraciles. Du point de vue évolutif, il y aurait donc eu d'abord apparition des désoxyribonucléotides, puis de l'ADN à uracile (ADN-U), puis d'ADN à thymine (ADN-T), qui se serait progressivement imposé. D'après Patrick Forterre, il est même probable que l'ADN-T ait été « inventé deux fois », chez des virus différents.
Les virus à ARN seraient ici des reliques du monde à ARN, les virus à ADN-U seraient alors des reliques du monde ayant précédé celui à ADN-T.
Les virus, premiers organismes à ADN
Les virus à ADN pourraient être plus anciens que la première cellule à ADN : la première cellule à ADN l'aurait donc emprunté à un ou plusieurs virus, sous la pression d'une course aux armements (théorie de la reine rouge).
Didier Raoult et Jean-Michel Claverie ont ainsi découvert le mimivirus : un virus géant à ADN (son génome étant deux fois plus long que le plus petit génome bactérien connu). La particularité de ce virus est qu'il peut produire des protéines impliquées dans la traduction de l'ARN en protéines (comme des enzymes chargeant des acides aminés sur des ARNt), il pourrait donc avoir pour ancêtres des virus plus anciens que la première cellule à ADN.
Eugène Koonin et ses collègues ont mis en avant, en comparant des génomes séquencés, que la plupart des enzymes impliqués dans la réplication de l'ADN sont différents entre les eubactéries et les eucaryotes (accompagnés des archées). Ils en concluent que l'ADN aurait été inventé indépendamment dans la lignée des eubactéries et celle conduisant aux eucaryotes et aux archées.
De même, les enzymes de réplications des virus à ADN sont très différentes d'un virus à l'autre, ainsi que par rapport aux enzymes cellulaires jouant le même rôle.
Ces indices laissent penser que les enzymes liées à l'ADN sont apparues au cours d'un « premier âge » du monde à ADN, où existaient cellules à ARN et virus à ARN et à ADN.
Passage de l'ADN dans les cellules
La nature du génome du plus ancien ancêtre commun à tous les êtres vivants (que les scientifiques prénomment LUCA) reste inconnue : faisait-il encore partie du monde à ARN, ou avait-il déjà un génome à ADN ? Quoi qu'il en soit, LUCA est le fruit d'une longue évolution. Le génome des premières cellules, qui ont précédé LUCA, était sans doute constitué par des molécules d'ARN et non pas d'ADN. Contrairement à l'ADN, l'ARN peut en effet jouer à la fois le rôle d'enzyme et de matériel génétique. Stanley Miller et Christian de Duve pensent que l'apparition de l'ARN a été elle-même un évènement tardif, en effet, cette molécule ne semble pas pouvoir être synthétisée par les méthodes simples de la chimie prébiotique. L'ARN aurait donc été précédé par des molécules dont nous ne connaîtrons sans doute jamais la nature exacte.
Ce qui amène les scientifiques à penser que l'ARN a précédé l'ADN résulte de ce constat : les trois grandes lignées du vivant ne partagent que le système de biosynthèse des protéines, alors qu'elles diffèrent sur le système de réplication de l'ADN.
Il existe alors plusieurs hypothèses pour le passage de l'ADN (d'origine viral) dans les cellules vivantes : soit ce passage s'est produit une seule fois, soit il a pu avoir lieu plusieurs fois, indépendamment dans les différentes lignées. Dans le premier cas, les premiers gènes des enzymes de réplications auraient donc été remplacés par la suite par ceux d'un autre virus, formant ainsi les trois lignées.
L'hypothèse de Patrick Forterre est que les trois lignées du vivant trouvent leurs sources dans le remplacement du génome à ARN par le génome à ADN de trois virus différents. On retrouve notamment ici l'hypothèse d'une origine virale du noyau des eucaryotes.
Les travaux de Carl Woese semblent appuyer cette hypothèse, en démontrant que la vitesse d'évolution des protéines semble avoir chuté au moment de l'apparition des trois lignées. Cette diminution serait due au passage de l'ARN à l'ADN, les génomes à ADN étant plus stables, et donc moins sensibles aux mutations.
Dans ce scénario, l'apparition de trois lignées uniquement s'explique par le fait que les cellules à ADN ont peu à peu supplanté les cellules à ARN, empêchant ainsi l'apparition de nouvelles lignées par passage ARN→ADN.
Autres modèles
Controverse sur la véracité de la théorie
On ne peut pas remonter le temps pour aller voir comment était l’océan primitif et comment il a évolué. Mais on peut trouver des témoignages indirects. Le passé laisse des traces dans le présent. Si on a les bons outils, théoriques et observationnels, on peut déduire le passé à partir du présent. Par exemple, les techniques autocatalytiques d’aujourd’hui se sont en quelque sorte fossilisées depuis des milliards d’années, puisqu’elles n’ont pas ou peu évolué. Elles nous renseignent donc sur un passé très lointain. En combinant ces informations avec d’autres, on peut espérer remonter encore plus loin dans le temps. Des expériences en laboratoire de chimie prébiotique peuvent apporter des renseignements précieux.
La théorie de la génération spontanée peut être combinée avec celle de la panspermie afin d'élargir le volume de matières mis en jeu à d'autres planètes viables. Néanmoins, elle rajoute la question de la probabilité que des organismes vivants puissent être véhiculés par des astéroïdes d'une planète viable à une autre au hasard des collisions intersidérales. Remarque, si l'on suppose que l'Univers contient des millions de planètes propices à la vie comme semblent l'indiquer les progrès de l'exploration spatiale, cette théorie mérite d'être étudiée.
Milieu de collision planète et noyaux de comètes
Selon des travaux de modélisation présentés en 2010, la compression brutale des matériaux constituant le noyau de glace d'une comète, au moment du choc avec une planète pourrait donner lieu à la production d'acides aminés. Cette modélisation a pris comme base 210 molécules diluées dans un mélange d'eau, de méthanol, d'ammoniac, de dioxyde de carbone et de monoxyde de carbone estimés couramment présentes dans le noyau des comètes. Avec une vitesse d'impact de 29 km/s, la pression atteint 10 gigapascals et une température de 700 K. Selon les auteurs, d'autres simulations, à pressions et températures plus élevées conduisent à des réactions chimiques encore plus complexes. Selon le modèle, une simulation de 47 gigapascals et une température de 3 141 K pour les 20 premières picosecondes de l'impact fait naître des molécules complexes, dont de grosses molécules à liaisons carbone-azote qui pourraient être les premières briques de la vie. Le calcul a nécessité environ (?) million d'heures d'ordinateur sur le superordinateur du cluster Atlas du Lawrence Livermore.
Sources hydrothermales : le monde du soufre
Les sources hydrothermales ont été découvertes en 1977 à 2 600 mètres de profondeur, là où deux plaques tectoniques se séparent.
Les monts hydrothermaux sont situés sur la couche sédimentaire. Leur diamètre à la base varie de 25 à 100 mètres et leur hauteur varie de 70 à 100 mètres. Les cheminées de ces fumeurs sont parfois recouvertes d'une croûte d'oxyde de manganèse. Les fumeurs situés sur ces monts sont composés d'un solide friable dont la couleur varie du gris noir à l'ocre, ce sont des sulfures de fer, de cuivre et de zinc.
Ces sources sont particulièrement intéressantes car on y a trouvé la vie où on la croyait impossible : milieu privé d'oxygène, à haute température, chargé de métaux et de soufre, dans l'obscurité la plus totale. Cependant les gradients de température importants autour de ces zones et le fait que les ultraviolets destructeurs ne parviennent pas si profondément (alors qu'ils détruisent toute molécule formée à la surface) sont de bonnes conditions pour l'apparition de la vie.
Ces organismes ont les mêmes formes que ceux que l'on connait plus près de la surface (ADN, protéines, sucres, etc.) mais puisent leur énergie de l'oxydation du sulfure d'hydrogène (H2S) pour transformer le carbone minéral en matière organique.
D'autre part, des expériences ont été menées, au laboratoire de géophysique de la Carnegie Institution de Washington, et ont montré que dans les conditions qui existent autour des évents, il y a formation d'ammoniac (NH3), forme réduite de l'azote qui est tant nécessaire à la formation des molécules organiques de la première partie et qui n'existait pas dans l'atmosphère oxydante. Les sources hydrothermales sont donc de bonnes sources de NH3.
Une origine extraterrestre primitive (exogenèse)
Une hypothèse alternative est que la vie se soit d'abord formée hors de la Terre.
Les composés organiques sont relativement fréquents dans l'espace, notamment dans les zones lointaines du système solaire où l'évaporation des composés volatils est très réduite. Certaines comètes présentent des couches externes de matière sombre, qu'on pense être une sorte de bitume formé par une combinaison de composés carbonés simples exposés aux rayons ultraviolets. La pluie de matériaux cométaires sur la Terre primitive pourrait avoir apporté des quantités de molécules organiques complexes, ce qui aurait favorisé l'apparition de la vie sur Terre. Ce serait plutôt là un apport de matériaux organiques, plus ou moins complexes (qui a dû se faire !), que l'apport de la vie elle-même.
Panspermie
Une hypothèse plus large est la panspermie : la vie même serait apparue « dans l'espace » puis disséminée sur Terre. Selon une variante, la vie serait apparue sur Mars d'abord et des impacts de comètes et d'astéroïdes sur Mars auraient projeté du matériel de la surface martienne sur Terre. Elle a quelques arguments en sa faveur :
l'environnement primitif de Mars était plus ou moins similaire à celui de la Terre primitive, et surtout a eu des conditions clémentes plus tôt que la Terre, car s'étant refroidi plus vite ;
les impacts majeurs sur Mars étaient moins susceptibles d'affecter gravement la totalité de la planète, bien qu'étant plus petite, car n'ayant pas eu d'océan global. Des impacts majeurs ont dû se produire en grand nombre durant l'Hadéen sur Terre, faisant bouillir une partie, voire la totalité, des océans ; la Terre a été périodiquement recouverte d'une atmosphère de vapeur brûlante (>100°C voire ~300°C) qui a pu stériliser (plusieurs fois) la planète entière ;
quelques éjectas (plus nombreux à ces époques reculées), ont pu faire le trajet Mars-Terre « assez rapidement », quelques années à quelques siècles, pour que d'hypothétiques spores puissent survivre à l'irradiation spatiale.
Il est encore plus difficile de trouver des indices pour justifier ces hypothèses que les théories plus classiques.
Ces théories d'une origine extraterrestre n'expliquent pas directement comment la vie est apparue, car elles ne font a priori que reporter le problème. Cependant, elles élargissent les conditions dans lesquelles la vie a pu apparaitre dans l'Univers. Les futurs échantillons de sols ramenés de Mars et de comètes permettront peut-être d'obtenir de nouveaux éléments de réponse.
Origine de l’évolution darwinienne plutôt qu’origine de la vie
Des scientifiques préfèrent étudier la question de l'origine de la vie à partir d’un paradigme ou d’un point de vue différent. Il s’agit de traiter la question de comment et quand l’évolution a commencé au lieu d’essayer de définir la vie et de délimiter une frontière quant à son origine.
Aujourd’hui il semble y avoir un consensus parmi les chercheurs sur l’origine de la vie (chimistes, géochimistes, biochimistes, exo/astrobiologistes, informaticiens, philosophes et historiens des sciences) qu’il est absolument nécessaire de trouver une définition universelle de la vie. Cependant le nombre de définitions ne cesse d’augmenter. De fait, la frontière entre systèmes vivants et systèmes non vivants de même que le moment à partir duquel le non vivant serait devenu vivant sont arbitraires.
On constate depuis quelques années un changement profond car le problème de la vie n’est plus de rechercher les principes qui la sous-tendent mais est devenu un problème de nature historique. La question n’est plus : « Qu’est-ce qui caractérise les organismes par rapport aux objets inanimés ? » ; mais plutôt : « comment ces caractéristiques se sont progressivement installées dans les systèmes que nous appelons des organismes? ». Finalement, la question deviendrait : « Comment l'évolution darwinienne a-t-elle émergé sur la Terre, il y a 4 milliards d'années environ, dans un monde qui ne la contenait pas encore? »
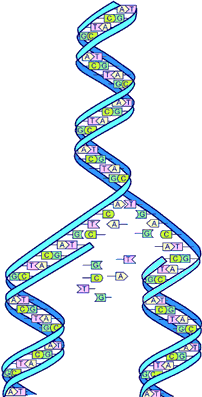
米勒-尤里实验
此表盘展示地质学时间及地球历史事件。冥古宙40亿前年部分为无生命时期,其余部分体现了生命之演进。最后二百万年的第四纪为人类时间,在图中太短而看不到。
在物质科学与无生源论中,****的研究对象主要是关于地球上的生命,如何经历约39到41亿年的演变,从无生物(或死物)成为生物。
演化论的解释
最早生命形式诞生 位在美国冰川国家公园的前寒武纪叠层石(stromatolite),可能是已知最早的生命化石纪录。 地球诞生时(后期重轰炸期后)的面貌和现在不同,包围在地球外表的水汽虽已凝结成液态性的水-海洋,但温度还是很高,那时具有活动力的火山遍布地表,不时喷出火山灰和岩浆,大气很稀薄,氢、一氧化碳等,各种气体于空中形成一朵朵的卷云,氧气很少,因无充足的大气层掩蔽,整个地球曝露在强烈的紫外线之下。此时云端的电离子不断引起风暴,而交加的雷电不时侵袭陆地。 遗传物质出现 几百年过去,这些物质越聚越多分子间互相影响,而形成更复杂的混合物,在这其间来自外太空的陨石也可能带来一些元素参与变化,RNA在生命最初的进化中扮演了一个重要角色。RNA比DNA的结构更加简单,而且是一种更加有效的化学催化剂。因此这就意味着RNA构成的生命比DNA生命更容易出现。DNA有两项特质:第一,它能通过转录产生mRNA,而mRNA则能够翻译出蛋白质,第二,它能自行复制,DNA这两项特质也是细菌类的有机生物的基本特质,而细菌是生命界最简单的生命体,也是目前我们可以找到最古老的化石。 DNA的复制本领来自其特殊的构造,DNA为双股螺旋,细胞的遗传消息都在上面,然而DNA在复制过程中也会出错,或是分子群的一小部分出错,如此复制工作就不尽完美,制造出来的蛋白质也可能完全不同,也就是如此演化便开始产生,一旦生命有了不同的型态,自然才能实施淘汰和选择的法则,生物才能一步步的演化下去,我们从化石中得知三十亿年前那些类似细菌的有机物之间,已有显著的不同。 时间比喻 若把地球诞生至今的这段日子当成一年,十一月的第三个星期鱼类才出现。
主要争议
概率问题 地球生物组织中的蛋白质具有高度的选择性,只使用了已发现氨基酸的不到五分之一,而这些氨基酸种类又是经过严格筛选出来的。按照随机产生的概率计算,即使生成一个很简单的具有生物活性的蛋白质,也许要远远大于可能性的次数才能随机产生,其自然概率也是小到几乎不可能出现。然而自然选择不是一个随机的过程,否则就不称其为选择了,因此这类质疑现在已经不成为主要争议。 原始大气成分 上述描述论及原始大气时,提到是一个缺氧的环境。这样地球就没有臭氧层的保护。在强烈的紫外线照射下,DNA,RNA及蛋白质难以存在。假如有氧气存在,其强烈的氧化能力也会破坏蛋白质等有机分子。 蛋白质和遗传物质的矛盾关系 上述解释在产生DNA/RNA遗传物质的时候只是一笔带过。然而在分子生物学研究发现,蛋白质和遗传物质存在严重的依赖关系。没有蛋白质的参与无法完成DNA/RNA的复制和组合,没有DNA/RNA携带的信息,蛋白质也就无从产生。 《新不列颠百科全书》说:「惟独这两种分子共同发挥作用,生物才可能在地上生存。」可是这套百科全书指出,这两种分子怎么会彼此紧密合作,「在****方面」仍然「是个关键性的哑谜」。而目前解释此一矛盾的理论,较为人所知的是「RNA世界学说」。