Les poissons sont des animaux vertébrés aquatiques à branchies, pourvus de nageoires et dont le corps est le plus souvent couvert d'écailles. On les trouve abondamment aussi bien dans les eaux douces que dans les mers : on trouve des espèces depuis les sources de montagnes (omble de fontaine, goujon) jusqu'au plus profond des océans (grandgousier, poisson-ogre). Leur répartition est toutefois très inégale : 50 % des poissons vivraient dans 17 % de la surface des océans (qui sont souvent aussi les plus surexploités).
Ils ont un rôle fondamental pour les hommes :
en tant que nourriture, partout dans le monde ; qu'ils soient pêchés dans la nature ou élevés en pisciculture ; ils sont aussi exploités à des fins récréatives, avec la pêche et l'aquariophilie, et sont parfois exposés dans de grands aquariums publics ; ils jouent et ont joué un rôle significatif dans de nombreuses cultures, en tant que déités et symboles religieux, ou sujets de contes, légendes, livres et films ; en jouant un rôle écosystémique important ; en contribuant aux cycles biogéochimiques, dont le cycle du carbone et en particulier en contribuant au puits de carbone océanique.
Dans la classification phylogénétique, obtenue par application des méthodes cladistiques, le groupe des poissons est paraphylétique. La classe des Poissons (Pisces) de la classification classique est obsolète. Les espèces actuelles (non-éteintes) de poissons sont réparties dans les taxons Actinoptérygiens (les plus communs), Chondrichtyens (les raies et requins), Dipneustes, Actinistiens, Myxinoïdes et Pétromyzontides.
Le premier inventaire ichtyologique de France semble être celui de Belon P., en 1555, intitulé La nature et la diversité des poissons avec leurs pourtraicts représentés au plus près du naturel.
Définition et classification
Définition
Un ptérois à antennes.
Le terme « poisson » est plus précisément employé pour désigner les crâniates non tétrapodes, c'est-à-dire des animaux avec un crâne cartilagineux ou osseux qui protège la partie antérieure du système nerveux, possédant des branchies toute leur vie et qui peuvent posséder des nageoires, mais pas de « pattes ». Les poissons ne forment pas un groupe phylogénétiquement homogène, à l'inverse des oiseaux ou des mammifères (voir plus bas).
Un poisson typique est « à sang froid » ; il possède un corps allongé lui permettant de nager rapidement ; il extrait le dioxygène de l'eau en utilisant ses branchies ou un organe respiratoire annexe lui permettant de respirer le dioxygène atmosphérique ; il possède deux paires de nageoires, les nageoires pelviennes et latérales, habituellement une ou deux (plus rarement trois) nageoires dorsales, une nageoire anale et une nageoire caudale; il possède une double mâchoire pour les gnathostomes et simple pour les agnathes ; il possède une peau généralement recouverte d'écailles ; ovipare, il pond des œufs et la fécondation peut être interne ou externe.
Il existe de nombreuses formes et tailles de poissons : ce dragon des mers, proche de l'hippocampe, se confond avec les algues grâces à ses nageoires.
Chacune de ces caractéristiques comporte toutefois des exceptions. Les thons, les espadons et certaines espèces de requins sont entre sang chaud et sang froid, et peuvent élever leur température corporelle au-dessus de celle de l'eau ambiante. Et de la même façon, la lampris-lune semble être un cas unique de poisson à sang chaud. La forme du corps et les performances natatoires varient considérablement, des nageurs très rapides capables de parcourir dix à vingt longueurs de leur corps par seconde (thons, saumons) aux poissons très lents mais mieux manœuvrants (comme les anguilles ou les raies) qui ne dépassent pas 0,5 longueur par seconde. Plusieurs groupes de poissons d'eau douce extraient le dioxygène de l'air comme de l'eau en utilisant des organes variés. Les dipneustes possèdent deux poumons similaires à ceux des tétrapodes ; les gouramis ont un « organe labyrinthe » qui fonctionne de la même manière ; les Corydoras extraient le dioxygène par l'estomac ou l'intestin. La forme du corps et la position des nageoires varient énormément, comme en témoigne la différence entre les hippocampes, les lophiiformes, les poissons globes ou les saccopharyngiformes. De même, la surface de la peau peut être nue (murènes) ou couverte d'écailles de différents types : placoïdes (requins et raies), cosmoïdes (cœlacanthes), ganoïdes, cycloïdes et cténoïdes. Certains poissons passent même davantage de temps hors de l'eau que dedans, comme les périophthalmes qui se nourrissent et interagissent entre eux sur des terrains boueux et ne retournent dans l'eau que pour se cacher dans leur terrier. Certaines espèces peuvent être ovovivipares ou vivipares.
La taille d'un poisson varie du requin baleine de 16 m au Schindleria brevipinguis d'à peine 8 mm.
Quelques espèces de poisson dulçaquicole disposent de poumons, si les Erythrinus de l'Amazone extraient 50 % du dioxygène dont ils ont besoin avec ceux-ci, les poumons sont l'unique moyen de respiration des Arapaima gigas ou des gymnotes.
Terminologie
Plusieurs types d'animaux aquatiques sont communément appelés « poissons » mais n'en sont pas vis-à-vis de la définition ci-avant.
Les termes relatifs aux poissons viennent de différentes racines :
Poisson se dit piscis (pisces au pluriel) en latin, d'où la pisciculture, piscivore, etc.
Poisson se dit ichthys en grec (plus exactement le mot grec ἰχθύς se transcrit ainsi en latin) ; l'Académie française a simplifié le groupe chth en cht en 1878 mais l'on retrouve le groupe complet en anglais par exemple ; d'où l'ichtyologie, Ichtus, l'ichtyocolle, etc.
Poisson se dit fish en anglais d'où le phishing.
Classification
À l'opposé des groupes tels que les oiseaux ou les mammifères, les poissons ne forment pas un clade : le groupe est paraphylétique, c'est-à-dire qu'il ne comporte pas tous les descendants de leur ancêtre commun. Pour cette raison, la « classe Pisces », comme on peut lire dans d'anciennes références, n'est plus utilisée en classification phylogénétique, chaque clade devant comporter tous les descendants du même ancêtre, ce qui amènerait à y adjoindre les Tétrapodes.
Les poissons sont classés dans les groupes principaux suivants (en grisé et précédés de l'obèle « † », les taxons éteints) :
Infra-embranchement des Agnathes ou Cyclostomes, (vertébrés sans mâchoires) Classe Hyperoartia (Hyperoartiens : poissons sans mâchoires) Ordre des Petromyzontiformes (lamproies) groupe des †Ostracodermes (terme qui regroupe les classes éteintes de poissons sans mâchoires) Classe †Pteraspidomorphi GOODRICH, 1909 (Ptéraspidomorphes, poissons primitifs sans mâchoires) Sous-classe †Heterostraci LANKESTER, 1868 (Hétérostracés, poissons primitifs sans mâchoires, Ordovicien → Dévonien) Sous-classe †Astrapida BERG, 1940 (Astrapides, poissons primitifs sans mâchoires, Ordovicien → Dévonien) Sous-classe †Arandaspida RITCHIE et GILBERT-THOMLINSON, 1977 (Arandaspides, poissons primitifs sans mâchoires, Ordovicien) Classe †Anaspida TRAQUAIR, 1899 (poissons primitifs sans mâchoires, Silurien → Dévonien supérieur) Classe †Thelodonti JAECKEL, 1911 (poissons primitifs sans mâchoires, Ordovicien → Dévonien) Classe †Cephalaspidomorphi GOODRICH, 1909 (Céphalaspidomorphes : poissons sans mâchoires à cuirasse articulée) Sous-classe †Osteostraci LANKESTER, 1868 (Ostéostracés : poissons à cuirasse ossifiée, Silurien moyen → Dévonien supérieur) Sous-classe †Galeaspida LIU, 1965 (Galéaspidés, poissons avec bouclier céphalique en forme de heaume, Silurien → Dévonien) Sous-classe †Pituriaspida (Pituriaspides, poissons cuirassés de "Pituri", Queensland, Australie, Dévonien moyen)
Classe Hyperoartia (Hyperoartiens : poissons sans mâchoires) Ordre des Petromyzontiformes (lamproies)
Ordre des Petromyzontiformes (lamproies)
groupe des †Ostracodermes (terme qui regroupe les classes éteintes de poissons sans mâchoires) Classe †Pteraspidomorphi GOODRICH, 1909 (Ptéraspidomorphes, poissons primitifs sans mâchoires) Sous-classe †Heterostraci LANKESTER, 1868 (Hétérostracés, poissons primitifs sans mâchoires, Ordovicien → Dévonien) Sous-classe †Astrapida BERG, 1940 (Astrapides, poissons primitifs sans mâchoires, Ordovicien → Dévonien) Sous-classe †Arandaspida RITCHIE et GILBERT-THOMLINSON, 1977 (Arandaspides, poissons primitifs sans mâchoires, Ordovicien) Classe †Anaspida TRAQUAIR, 1899 (poissons primitifs sans mâchoires, Silurien → Dévonien supérieur) Classe †Thelodonti JAECKEL, 1911 (poissons primitifs sans mâchoires, Ordovicien → Dévonien) Classe †Cephalaspidomorphi GOODRICH, 1909 (Céphalaspidomorphes : poissons sans mâchoires à cuirasse articulée) Sous-classe †Osteostraci LANKESTER, 1868 (Ostéostracés : poissons à cuirasse ossifiée, Silurien moyen → Dévonien supérieur) Sous-classe †Galeaspida LIU, 1965 (Galéaspidés, poissons avec bouclier céphalique en forme de heaume, Silurien → Dévonien) Sous-classe †Pituriaspida (Pituriaspides, poissons cuirassés de "Pituri", Queensland, Australie, Dévonien moyen)
Classe †Pteraspidomorphi GOODRICH, 1909 (Ptéraspidomorphes, poissons primitifs sans mâchoires) Sous-classe †Heterostraci LANKESTER, 1868 (Hétérostracés, poissons primitifs sans mâchoires, Ordovicien → Dévonien) Sous-classe †Astrapida BERG, 1940 (Astrapides, poissons primitifs sans mâchoires, Ordovicien → Dévonien) Sous-classe †Arandaspida RITCHIE et GILBERT-THOMLINSON, 1977 (Arandaspides, poissons primitifs sans mâchoires, Ordovicien)
Sous-classe †Heterostraci LANKESTER, 1868 (Hétérostracés, poissons primitifs sans mâchoires, Ordovicien → Dévonien)
Sous-classe †Astrapida BERG, 1940 (Astrapides, poissons primitifs sans mâchoires, Ordovicien → Dévonien)
Sous-classe †Arandaspida RITCHIE et GILBERT-THOMLINSON, 1977 (Arandaspides, poissons primitifs sans mâchoires, Ordovicien)
Classe †Anaspida TRAQUAIR, 1899 (poissons primitifs sans mâchoires, Silurien → Dévonien supérieur)
Classe †Thelodonti JAECKEL, 1911 (poissons primitifs sans mâchoires, Ordovicien → Dévonien)
Classe †Cephalaspidomorphi GOODRICH, 1909 (Céphalaspidomorphes : poissons sans mâchoires à cuirasse articulée) Sous-classe †Osteostraci LANKESTER, 1868 (Ostéostracés : poissons à cuirasse ossifiée, Silurien moyen → Dévonien supérieur) Sous-classe †Galeaspida LIU, 1965 (Galéaspidés, poissons avec bouclier céphalique en forme de heaume, Silurien → Dévonien) Sous-classe †Pituriaspida (Pituriaspides, poissons cuirassés de "Pituri", Queensland, Australie, Dévonien moyen)
Sous-classe †Osteostraci LANKESTER, 1868 (Ostéostracés : poissons à cuirasse ossifiée, Silurien moyen → Dévonien supérieur)
Sous-classe †Galeaspida LIU, 1965 (Galéaspidés, poissons avec bouclier céphalique en forme de heaume, Silurien → Dévonien)
Sous-classe †Pituriaspida (Pituriaspides, poissons cuirassés de "Pituri", Queensland, Australie, Dévonien moyen)
Infra-embranchement des Gnathostomes (vertébrés à mâchoires) Classe †Placodermi (Placodermes : poissons à cuirasse articulée) Classe †Acanthodii (Acanthodiens : requins épineux) Classe Chondrichthyes (Chondrichtyens : poissons cartilagineux : requins et les raies) Superclasse Osteichthyes (Ostéichthyens : poissons osseux) Classe Actinopterygii (Actinoptérygiens : poissons à nageoires rayonnées) Classe Sarcopterygii (Sarcoptérygiens : poissons à nageoires charnues) Sous-classe Coelacanthimorpha (cœlacanthes) Sous-classe Dipnoi (dipneustes)
Classe †Placodermi (Placodermes : poissons à cuirasse articulée)
Classe †Acanthodii (Acanthodiens : requins épineux)
Classe Chondrichthyes (Chondrichtyens : poissons cartilagineux : requins et les raies)
Superclasse Osteichthyes (Ostéichthyens : poissons osseux) Classe Actinopterygii (Actinoptérygiens : poissons à nageoires rayonnées) Classe Sarcopterygii (Sarcoptérygiens : poissons à nageoires charnues) Sous-classe Coelacanthimorpha (cœlacanthes) Sous-classe Dipnoi (dipneustes)
Classe Actinopterygii (Actinoptérygiens : poissons à nageoires rayonnées)
Classe Sarcopterygii (Sarcoptérygiens : poissons à nageoires charnues) Sous-classe Coelacanthimorpha (cœlacanthes) Sous-classe Dipnoi (dipneustes)
Sous-classe Coelacanthimorpha (cœlacanthes)
Sous-classe Dipnoi (dipneustes)
Certains paléontologues considèrent que les conodontes sont des chordés et les considèrent comme des poissons primitifs ; voir l'article Vertébré.
Les différents groupes de poissons pris tous ensemble comprennent plus de la moitié des vertébrés connus. Il y a près de 28 000 espèces de poissons existantes (sans compter les espèces disparues), dont près de 27 000 poissons osseux, le reste étant formé d'environ 970 requins, raies et chimères et environ 108 lamproies et myxines. Un tiers de toutes ces espèces est renfermé dans les neuf plus grandes familles, qui sont (des plus grandes aux plus petites) : Cyprinidae, Gobiidae, Cichlidae, Characidae, Loricariidae, Balitoridae, Serranidae, Labridae, et Scorpaenidae. D'un autre côté, environ ** familles sont monotypiques (ne contiennent qu'un seul genre, parfois monospécifique). On prédit que le nombre total d'espèces de poissons en existence serait de 32 500.
Génomique
Les poissons contemporains sont les vertébrés chez lesquels ont observe les plus grands et les plus petits génomes (parmi les vertébrés), phénomène qui a une « signification évolutive » encore incomprise. Le génome est plus petit chez les poissons à nageoires à rayons que chez les poissons cartilagineux, hormis chez les polyploïdes (qui explique en grande partie les variations de taille de génome au sein de ces deux groupes). Le génome des poissons d'eau douce (et eurybiontes) est plus grands que celui des espèces apparentées marines et sténobiontes. Les différences de taille de génome ne semblent pas liées au taux métabolique propre à l'espèce mais elles sont positivement corrélée avec la taille des œufs, ce qui peut faire évoquer un lien avec l'évolution des soins parentaux.
Évolution
Haikouichthys
Dans un contexte évolutif, l'ensemble des taxons désignés par le terme poisson n'est plus considéré comme tel, ces taxons ayant des histoires évolutives différentes et formant donc différents clades.
Certains pensent que les poissons ont évolué à partir d'une créature du type ascidie (dont les larves ont des ressemblances avec les poissons primitifs) ; les premiers ancêtres des poissons auraient alors conservé leur forme larvaire à l'état adulte par néoténie, mais l'inverse est aussi possible. Les fossiles candidats au statut de « premier poisson » connus sont Haikouichthys, Myllokunmingia et Pikaia.
Les tout premiers fossiles de poissons ne sont guère nombreux, ni de bonne qualité : peut-être les poissons primitifs étaient-ils rares ou mal fossilisables ou les conditions taphonomiques mauvaises. Cependant, le poisson devint une des formes de vie dominantes du milieu aquatique et a donné naissance aux branches évolutives menant aux vertébrés terrestres comme les amphibiens, les reptiles et les mammifères.
L'apparition d'une mâchoire articulée semble être la raison majeure de la prolifération ultérieure des poissons, car le nombre d'espèces de poissons agnathes devint très faible. Les premières mâchoires ont été trouvées dans les fossiles de placodermes. On ignore si le fait de posséder une mâchoire articulée procure un avantage, par exemple pour la préhension ou la respiration.
Les poissons ont aussi coévolué avec d'autres espèces (prédateurs, pathogènes et parasites notamment, mais aussi parfois des espèces symbiotes). Durant leurs migrations (longues et sur de longues distances pour les saumons et lamproies, et plus encore pour les anguilles), ils peuvent transporter (dispersion) un certain nombre de propagules d'autres organismes (ectoparasitisme, endozoochorie, œufs viables non digérés….)
Vers nageurs
Branchiostoma lanceolatum
La fonctionnalité essentielle qui a initialement structuré cet ordre a été la capacité de nager dans l'eau (ce que ne savaient faire ni les mollusques primitifs, ni les arthropodes primitifs). Toutefois, cette capacité n'a pas conduit à une explosion radiative : par elle-même, elle ne donne pas une autonomie fonctionnelle suffisante pour que les organismes puissent se spécialiser de manière très libre.
L'histoire de cette lignée paraît laborieuse : la découverte progressive de la tête et de la mâchoire, puis l'exploration des membres jusqu'au stade tétrapode, et enfin, la conquête de l'environnement aérien, aboutissant à l'explosion radiative des sauriens.
Agnathes
Une lamproie marine.
Les agnathes regroupent des animaux à corde dorsale et à crâne, mais sans mâchoires. Leur vie en milieu aquatique les a fait longtemps classer parmi les poissons.
Le taxon n'est pas considéré comme monophylétique, et doit être considéré comme obsolète dans une classification phylogénétique. Il comprend :
les myxines, qui ont finalement été séparées des vertébrés ;
les lamproies, qui y restent rattachées.
Les myxines et lamproies partagent des caractères morphologiques ancestraux à tous les crâniates, qui seront perdus chez les gnathostomes. Leur bouche rudimentaire, qui se comporte comme une ventouse, ne possède pas de mâchoires, et ne peut donc pas modifier son ouverture. Leur squelette est cartilagineux et composé d'une capsule crânienne et d'une colonne vertébrale sans côtes.
Si les agnathes actuels sont peu nombreux, de nombreux fossiles d'agnathes sont présents dans les sédiments du paléozoïque. Les agnathes furent les tout premiers crâniates à apparaître.
Les fonctionnalités explorées à ce stade évolutif portent sur le fonctionnement de la tête :
la céphalisation, commune chez tous les vermiformes, s'accentue et le cerveau s'entoure d'un crâne ;
des branchies apparaissent, de forme variable suivant les groupes ;
la bouche se dote de dents.
Poissons cartilagineux (Chondrichtyens)
Un grand requin blanc de 3,5 mètres environ.
Chez les Chondrichthyens, aussi appelés « poissons cartilagineux », il n'y a globalement pas d'ossification endochondrale. Le squelette est donc très majoritairement composé de cartilage, et pas d'« os vrai ». On peut y trouver les différentes espèces de requins, de raies et de chimères. Il faut ajouter que les « os vrais » peuvent tout de même être observés chez les Chondrichtyens, mais en petite quantité. La généralisation de l'os enchondral ne se trouvera que chez les ostéichtyens, étant par ailleurs leur synapomorphie principale.
Les principales synapomorphies des chondrichthyens incluent une couche de cartilage calcifié prismatique et, chez les mâles, les nageoires pelviennes portent des claspers pelviens (organes servant à l'accouplement).
Ces poissons bénéficient de deux autres acquis qui leur permettent d'être des nageurs plus performants et réactifs : l'oreille interne qui se perfectionne et acquiert un troisième canal semi-circulaire, et surtout les fibres nerveuses qui se couvrent de myéline, ce qui permet une transmission plus rapide de l'influx nerveux.
Poissons osseux (Ostéichtyens au sens classique)
Un poisson mandarin, un exemple de poisson osseux.
Comme leur nom l'indique, l'innovation la plus notable des poissons osseux est l'os. Le tissu osseux périchondral qui renforçait certains cartilages se généralise, et conduit à deux types d'os d'origines différentes :
l'os enchondral (associé au mésoderme), qui remplace au cours du développement les pièces cartilagineuses du squelette interne ;
surtout, l'os dermique, qui se forme à partir du derme (d'origine mésodermique), apparaît. Il donne les os de la boîte crânienne et les ceintures scapulaires, ainsi que les rayons des nageoires (qui évolueront ultérieurement en membres).
On observe aussi la présence de sacs aériens connectés au tube digestif qui donneront les poumons des vertébrés terrestres et les vessies natatoires des Actinoptérygiens. Ces sacs aériens sont soupçonnés chez certains Gnathostomes fossiles. Les tentatives d'émancipation du milieu aquatique seraient alors apparues dans ce clade.
Les principales fonctionnalités évolutives explorées au niveau des poissons osseux sont l'articulation de la mâchoire, de plus en plus structurée, et la forme et la mobilité des nageoires.
Poisson à nageoires charnues
Dipneuste, dessin de Heinrich Harder (autour de 1920)
On peut par exemple compter comme poissons à nageoires charnues (sarcoptérygiens) les cœlacanthes, les dipneustes ou encore les tétrapodes. Plusieurs innovations se retrouvent dans ce groupe :
le membre monobasal : l'attache basale se réduit à un seul élément (fémur, omoplate et humérus) et des muscles permettent l'articulation indépendante des rayons (doigts) ;
les dents se couvrent d'émail.
Il apparaît donc évident, vu les membres de ce groupe, que pour rendre le groupe des poissons comme monophylétique (et donc le considérer en biologie, voir cladistique), il faut notamment y inclure tous les vertébrés terrestres (dont les humains font évidemment partie).
Morphologie et anatomie
Morphologie
Aspect général
Anatomie d'un poisson à nageoires rayonnées A Nageoire dorsale B Rayon C Ligne latérale D Rein E Vessie natatoire F Appareil de Weber G Oreille interne H Cerveau I Narine L Orbite M Branchie N Cœur O Estomac P Vésicule biliaire Q Rate R Gonades S Nageoire ventrale ou pelvienne (par paire) T Colonne vertébrale U Nageoire anale V Nageoire caudale.
Schéma du Lampanyctodes hectoris 1 Opercule 2 Ligne latérale 3 Nageoire dorsale 4 Nageoire molle ou adipeuse 5 Pédoncule caudal ou queue 6 Nageoire caudale 7 Nageoire anale 8 Photophores 9 Nageoire ventrale ou pelvienne (par paire) 10 Nageoire pectorale (par paire)
Épiderme
Chez les vertébrés aquatiques : (Agnathes, poissons, larves d'amphibien) :
Les cellules épidermiques sont toutes vivantes, éliminations sans modification elles tombent. Peu épais (5 -9 couches cellulaires), il y a des échanges osmotiques et ioniques.
Nageoires
Nageoire dorsale d'un Chevesne.
La plupart des poissons se déplacent en contractant alternativement les muscles insérés de chaque côté de la colonne vertébrale. Ces contractions font onduler le corps de la tête vers la queue. Lorsque chaque ondulation atteint la nageoire caudale, la force propulsive créée pousse le poisson vers l'avant.
Les nageoires du poisson sont utilisées comme stabilisateurs. La nageoire caudale sert aussi à augmenter la surface de la queue, augmentant ainsi la poussée lors de la nage, et donc la vitesse. Le corps fuselé des poissons permet de diminuer les frictions lorsqu'ils nagent, et donc d'éviter qu'ils soient ralentis par la résistance de l'eau. De plus, leurs écailles sont enrobées d'un mucus qui diminue les frottements.
Anatomie
Comportement
Comportement social
De nombreuses espèces de poissons, tels les demoiselles (Pomacentridés), les poissons zèbres (Cyprinidés) ou les néons (Characidés), etc. ont un instinct grégaire et préfèrent vivre en bancs. D'autres, tels les requins, sont plutôt solitaires. Certains sont même si agressifs que la rencontre d'un congénère peut entraîner la mort de l'un d'eux après une rude bataille : c'est le cas du combattant du Siam (Anabantidés).
Migration des poissons
Menaces et conservation
Surpêche
Pour les poissons comestibles comme la morue et le thon, la menace principale est la surpêche. Lorsque la surpêche persiste, elle finit par causer une diminution de la population de poissons (le « stock ») car les individus ne peuvent pas se reproduire assez vite pour compenser la perte due à la pêche. Un exemple de surpêche catastrophique très étudié est celui de la sardine du Pacifique (Sadinops sagax caerulues), qui était pêchée près des côtes de Californie. Le maximum avait été atteint en 1937 avec 790 000 tonnes, puis la quantité pêchée a décru pour atteindre à peine 24 000 t en 1968, date à laquelle cette industrie s'arrêta faute d'être rentable. Une telle « extinction commerciale » ne veut pas dire que l'espèce elle-même est éteinte, mais seulement qu'elle n'est plus économiquement viable. La pêche minotière a également localement conduit à la surpêche.
La principale source de tension entre l'industrie de la pêche et la science halieutique est la recherche d'un équilibre entre la conservation des espèces pêchées, et la préservation du revenu des pêcheurs. Dans des zones comme l'Écosse, Terre-Neuve ou l'Alaska, où l'industrie des pêches est le principal employeur, le gouvernement est particulièrement impliqué dans cet équilibre, en maintenant à la fois un stock suffisant et des ressources suffisantes pour les pêcheurs. D'un autre côté, les scientifiques promeuvent une protection toujours accrue pour les stocks, en prévenant que de nombreux stocks pourraient disparaitre dans les cinquante prochaines années.
Selon le WWF, « 80 % des stocks mondiaux de poissons utilisés à des fins commerciales sont déjà surpêchés ou menacés de l’être. Par ailleurs, 40 % de tous les animaux marins capturés finissent comme prises accessoires et sont rejetés morts ou moribonds par-dessus bord. Et comme le poisson d’élevage est le plus souvent nourri avec de l’huile de poisson et/ou de la farine de poisson, les élevages contribuent également au pillage des mers ».
Guide des poissons gravement menacés d'extinction, à éviter à tout prix à la consommation
Guide des poissons non recommandables à la consommation
Guide des poissons acceptables à la consommation
Guide des poissons à privilégier à la consommation par rapport à d'autres
Pollution de l'eau, des sédiments et destruction des habitats
Les poissons sont notamment vulnérables à de nombreux polluants (plomb, mercure et autres métaux, insecticides... ) et à des problèmes de féiminisation (imposex) induits par des perturbateurs endocriniens.
Une des menaces sur les écosystèmes (marins et d'eau douce) est la dégradation physique, chimique et écologique des habitats ; celle-ci est causée par la pollution de l'eau, la construction de grands barrages, le réchauffement, l'eutrophisation, l'acidification et la baisse du niveau d'eau par les activités humaines, et doivent faire face à la concurrence et aux pathogènes d'espèces introduites. Un exemple de poisson en danger à cause d'un habitat modifié est l'esturgeon Scaphirhynchus albus, vivant dans les cours d'eau en Amérique du Nord, ceux-ci ayant été modifiés de différentes manières.
Espèces exotiques envahissantes
L'introduction d'espèces exotiques dont beaucoup sont devenues invasives s'est produite à de nombreux endroits et pour de nombreuses raisons, dont le ballastage des navires de commerce. Un exemple bien connu et étudié est l'introduction de la perche du Nil dans le lac Victoria. À partir des années 1960, la perche du Nil introduite pour la pêche a progressivement exterminé les 500 espèces de cichlidés que l'on ne trouvait nulle part ailleurs que dans ce lac ; certaines espèces ne survivent que grâce à des programmes de reproduction en captivité, mais d'autres sont probablement éteintes. Parmi les espèces de poissons invasives ayant causé des problèmes écologiques, on peut noter les carpes, les tête-de-serpent, les tilapias, la perche européenne, la truite fario, la truite arc-en-ciel ou la lamproie marine.
Conservation
Le requin-baleine, le plus grand poisson au monde, est classé comme « vulnérable ».
Statut de préservation
En 2006, la liste rouge de l'UICN comprenait 1 173 espèces de poissons menacées d'extinction. Cette liste incluait des espèces comme la morue de l'Atlantique, le Cyprinodon diabolis, les cœlacanthes ou le grand requin blanc. Comme les poissons vivent sous l'eau, ils sont plus compliqués à étudier que les animaux terrestres ou les plantes, et on manque toujours d'informations sur les populations de poissons. Les poissons d'eau douce semblent particulièrement menacés, car ils vivent souvent dans des zones restreintes.
Contribution des poissons aux puits de carbone
À partir du dioxyde de carbone dissous dans l'eau, les poissons marins produisent constamment dans leur intestin des carbonates peu solubles.
Par exemple, le flet européen synthétise et rejette chaque heure et en moyenne 18 micromoles de carbone par kg de poisson (sous forme de calcite). Les poissons contribueraient ainsi de 3 % à 15 % du puits de carbone océanique (voire 45 % si l'on prenait les hypothèses les plus « optimistes »). De plus, les poissons ont une marge de tolérance à la température et un climat chaud associé à la surpêche tendent à réduire le nombre de grands poissons ; or, une petite taille du poisson et une eau plus chaude favoriseraient cette formation de carbonates de calcium ou de magnésium (qui sont éliminés avec la nécromasse, les fèces ou des boulettes de mucus).
Hélas, ces carbonates plus riches en magnésium sont aussi plus solubles à grande profondeur. Ils peuvent alors relarguer une partie de leur carbone, mais en tamponnant le milieu, au point que cela pourrait expliquer jusqu'à un quart de l'augmentation de l'alcalinité titrable des eaux marines dans les 1 000 mètres sous la surface (cette anomalie de dureté de l'eau était jusqu'ici controversée car non expliquée par les océanographes).
Hélas encore, c'est aussi dans les zones les plus favorables à ce piégeage du carbone (plateaux continentaux où se concentre environ 80 % de la biomasse en poisson) que la surpêche est la plus intense et que les zones mortes ont fait disparaître le plus de poissons.
Mesures de protection
On cherche à établir des indicateurs ou indices de biodiversité en poissons, notamment pour les milieux d'eau douce, lacs et mares y compris.
Les échelles à poissons
Le poisson dans la culture
Dans les récits fondateurs
Gravure préhistorique de l'époque néolithique, montrant une silhouette mi-homme mi-poisson et de nombreux poissons.
Dans la mythologie égyptienne
Le tilapia, poisson qui a coutume de mettre ses petits dans sa bouche en cas de danger et de les laisser ressortir une fois la menace passée, est associé à la déesse Hathor, emblème de la résurrection.
Lorsque Seth découpa le corps d'Osiris en 14 morceaux et les dispersa dans le monde, seuls 13 furent retrouvés. Le 14 (le sexe) avait été dévoré par un poisson.
Dans la mythologie gréco-romaine
Triton est une divinité marine, fils de Poséidon et d'Amphitrite. Il est figuré avec une tête et un buste d'homme et une queue de poisson.
Dans la mythologie celtique
Dans le judéo-christianisme
Selon la tradition juive, Dieu servira aux justes Léviathan, poisson monstrueux, comme nourriture à la fin des temps.
Jonas, selon la Bible (Jonas, 2:1), a passé trois jours et trois nuits dans le ventre d'un poisson : « Yahvé fit qu'il y eut un grand poisson pour engloutir Jonas. Jonas demeura dans les entrailles du poisson trois jours et trois nuits. »
Après sa résurrection, Jésus, sur les bords du lac de Tibériade, prépare sur la braise un repas de poissons pour les disciples. Déjà lors des multiplications des pains, il avait fait distribuer du poisson aux foules affamées.
Très tôt, le poisson deviendra un symbole majeur du christianisme primitif.
Le poisson est devenu pour les catholiques le repas du vendredi, mais aussi le mets de pénitence et du temps de carême, ainsi que la nourriture ordinaire des religieux.
Dans les légendes du Moyen Âge
Les sirènes était des êtres mi-femme mi-poisson.
La constellation des Poissons
Dans les réalisations artistiques
La stalle du vendeur de poissons, huile sur toile attribuée à Arnout de Muyser (XVI siècle).
En dessin, peinture, mosaïque, sculpture
Le poisson rouge de Paul Klee
Les poissons rouges de Henri Matisse
Le poisson de Joan Miro
Le poisson volant de Calder
Mulet gris et camélias, estampe de Hiroshige.
Dans diverses cultures, de l'Antiquité à nos jours, le poisson a été l'objet de très nombreuses œuvres.
En musique
La Truite de Franz Schubert.
Chansons : « la Maman des poissons » et « le Poisson Fa » de Boby Lapointe.
Au cinéma
Gang de requins, film d'animation.
Le Cauchemar de Darwin, film documentaire portant sur la pêche de la perche du Nil dans le Lac Victoria.
Le Monde de Nemo, film d'animation avec comme héros un poisson clown.
Les dents de la mer, film dramatique et de suspens dans la station balnéaire d'Amity
En poésie
« Sardines à l'huile » et « Les poissons mélomanes » poèmes issus des « Petites élégies falotes » du recueil « La négresse blonde » de Georges Fourest.
Dans la bande dessinée
Quelques records
Taille
Le paiche d'Amazonie, ou Arapaïma, est le plus gros poisson dulçaquicole : il peut atteindre 4,5 m et 200 kg. Le plus gros poisson connu est le requin baleine (16 m, 10 t).
Selon les Proceedings of the Royal Society (janvier 2006), le plus petit poisson serait Paedocypris progenetica, un cyprinidé d'eau douce découvert par Maurice Kottelat et Tan Heok Hui, du Raffles Museum (Singapour). La femelle Paedocypris est mature à partir de 7,9 mm de long. Adultes, les femelles mesurent 10,3 mm et les mâles, 11,4 mm. Il détrône un gobie nain de l'océan Indien décrit en 1981, Trimmatom nanus, adulte à 8 mm. Un poisson du même genre, Paedocypris micromethes, légèrement plus grand (femelle adulte à partir de 8,8 mm), a été identifié à Sarawak. Ces poissons vivent dans les forêts marécageuses constituées d'arbres inondés poussant sur un sol de tourbe détrempée, mou et épais de plusieurs mètres. L'eau y est rouge sombre et très acide.
Selon une équipe de l'université de Washington, Photocorynus spiniceps est le plus petit vertébré connu : le mâle de cette espèce de baudroie de la famille des Cérates, découverte dans les abysses du large des Philippines, long de seulement 6,2 mm, vit en parasite sur le dos d'une femelle mesurant 46 mm. Celle-ci pourvoit aux besoins en nourriture d'un mâle ne se limitant pratiquement qu'à un appareil reproducteur (Pietsch et al., Ichtyological Research, 2005).
Si le mâle Photocorynus spiniceps est plus petit, notons qu'en moyenne Paedocypris progenetica détient le record du plus petit poisson connu.
Puissance
Dunkleosteus terrelli, un poisson marin à plaques cuirassées qui vivait il y a 400 Ma, mesurait jusqu'à 11 m de long et pouvait peser jusqu'à quatre tonnes. Après en avoir reconstitué la musculature, des scientifiques américains ont découvert en 2006 que les mâchoires de ce poisson étaient capables d'exercer une pression de 5 500 kg/cm, soit à peu près deux fois celle de l'actuel requin blanc et autant que la mâchoire d'un Tyrannosaurus. De surcroît, on estime que Dunkleosteus terrelli était capable d'ouvrir et de refermer sa gueule en un cinquantième de seconde (Philip Anderson et Mark Westneat, 2006).
 词典释义:
词典释义:
 ,
,  肉,
肉,  状物
状物

 罐头
罐头 得水
得水 肉
肉 群
群

 用的餐具
用的餐具 鳍
鳍 座
座 内脏
内脏
 在
在 钩上挣扎
钩上挣扎 ;
; , 鲛;
, 鲛;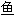 ;
; ,章
,章 ;
; ;
; ;
; ;
; , 蠹
, 蠹
 座
座 ;
;  状物; 猎
状物; 猎 具
具 , 淡水
, 淡水
 座
座 具
具
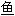
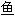 ; 鳜
; 鳜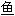

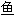
 ; 六须鲶
; 六须鲶 骨形天线
骨形天线 加工船
加工船 胶
胶 粉
粉 骨刺”状
骨刺”状 粉
粉 群探测[器、仪]
群探测[器、仪] ; 衣
; 衣
 甲板
甲板 翔脉
翔脉 装卸码头
装卸码头 回收
回收 口形缝合术
口形缝合术