 词典释义:
词典释义:

 年新生, 长新
年新生, 长新 子;
子; 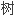
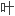
 新生期
新生期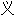 词:
词: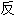
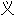 词:
词: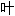 子,
子,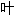 丛;
丛;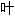 子;
子;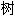 皮,茎皮;
皮,茎皮;_D._Don_(pabloendemico).jpg)
Feuilles d'un hêtre.
La feuille est, en morphologie végétale, l'organe spécialisé dans la photosynthèse chez les végétaux supérieurs. Elle est insérée sur les tiges des plantes au niveau des nœuds. À l'aisselle de la feuille se trouve un bourgeon axillaire. C'est aussi le siège de la respiration et de la transpiration. Les feuilles peuvent se spécialiser, notamment pour stocker des éléments nutritifs et de l'eau.
Pour accomplir son rôle, une feuille est généralement formée d'une lame plate et fine aérienne, le limbe, qui lui permet d'exposer à la lumière un maximum de surface. Mais il existe aussi des feuilles transformées, pour lesquelles le limbe est très réduit en ne joue plus de rôle photosynthétique ; elles sont transformées en vrilles, cataphylles, écailles sur les bourgeons, aériens (épines, aiguilles de conifères) ou souterrains (comme dans les bulbes, cormes), feuilles succulentes. C'est le parenchyme palissadique, un type particulier de tissu de la feuille, qui effectue la photosynthèse grâce à ses cellules contenant les chloroplastes, et donne à la feuille sa couleur verte. La feuille présente une grande variété de formes, de tailles, de teintes, de textures ou encore d'ornementations dans le règne végétal. Ces particularités de la feuille sont souvent caractéristiques d'une espèce végétale, ou au moins d'un genre.
Les feuilles de certains légumes, tel le navet, sont appelés « fanes » ; d'autres feuilles comestibles sont des brèdes.*
La feuillaison ou foliaison est un nom indénombrable qui désigne, pour les végétaux non persistants, l'apparition des feuilles, phase saisonnière concomitante au débourrement.
Description
La figure 1 montre les différentes parties de la feuille : un limbe plan (a) parcouru de nervures (b), avec souvent un pétiole (c) qui rattache la feuille à la tige, parfois élargi en gaine (d). Celle-ci peut « embrasser » la tige comme chez les poacées. Le pétiole peut être absent, la feuille est alors dite sessile. Il peut parfois être ailé, ou muni à sa base de stipules plus ou moins développés. Au point d'insertion du pétiole et de la tige, se trouve un bourgeon axillaire.
À la différence du reste de l'appareil végétatif de la plante (racine et tige), la feuille présente en général une symétrie bilatérale et non axiale.
Critères de description
Quatre critères permettent de décrire la feuille.
Composition
La feuille est composée de pectine, de cellulose et de lignine. Ces composants sont de grandes molécules chimiques « emprisonnant » de nombreux éléments minéraux tels que calcium, potassium, sodium, magnésium, soufre, phosphore. Lors de la décomposition des feuilles en humus, ces éléments sont relâchés dans le sol et contribuent à son amélioration.
Formes
La feuille est dite simple si le limbe est entier, ou composée s'il est découpé en plusieurs petites feuilles : les folioles. Selon la disposition des folioles sur l'axe principal de la feuille ou rachis, il est dit que la feuille est :
pennée, si les folioles sont disposées comme les barbes d'une plume ; imparipennée si leur nombre est impair (il y a une foliole terminale) ; paripennée dans le cas contraire ;
imparipennée si leur nombre est impair (il y a une foliole terminale) ;
paripennée dans le cas contraire ;
trifoliolée s'il y a trois folioles (trèfle) ;
palmée ou digitée si elles sont disposées comme les doigts de la main. Cependant les palmes, feuilles du palmier, qui sont en fait des feuilles entières, mais déchirées, ont parfois une forme pennée en forme de plume (dattier), ou encore costapalmée (Sabal palmetto) si la structure est intermédiaire entre les deux autres formes (soit l'aspect général d'un éventail mais avec un axe central assez marqué);
pédalée si chaque foliole est insérée sur la foliole voisine (hellébore) ;
La feuille peut être doublement composée ;
bipennée si les folioles sont composées de foliolules ;
tripennée si les folioles sont elles-mêmes composées.
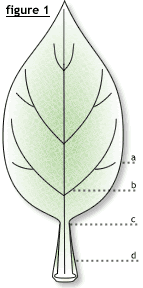
La forme de la feuille (fig. 2) : cordiforme, en forme de cœur (a), ovale (b) ou lancéolée (c), ou palmée, en forme de paume. Les formes sont très diversifiées : falciforme (en forme de faux), auriculée (avec des oreillettes à la base du limbe : laiteron), sagittée (en forme de flèche), peltée (lorsque le pétiole s'insère au milieu du limbe : capucine), décurrente lorsque le limbe se prolonge sur la tige par des ailes, perfoliée quand les oreillettes se soudent donnant l'impression que la tige traverse le limbe, connée quand deux feuilles opposées sont soudées par le limbe (chèvrefeuille)….
Division du limbe
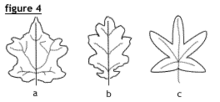
Le limbe peut être uni ou entier (comme fig. 1 et 2) ou plus ou moins profondément découpé (fig. 4). Dans ce dernier cas (fig.4), la feuille peut être :
lobée (a), lorsque les lobes sont peu profonds et atteignent environ 1/8 de la largeur de la feuille, pennatilobée (b), lorsque les lobes sont disposés en peigne, palmatilobée lorsque les lobes ont une disposition palmée.
pennatifide ou palmatifide, lorsque les divisions atteignent 1/4 de la largeur de la feuille et en tout cas moins de la moitié de celle-ci ;
pennatipartite (c), ou palmatipartite lorsque les découpures dépassent le milieu de la feuille ;
pennatiséquée (synonyme de composé pennée) ou palmatiséquée (composée palmée) si les divisions atteignent le rachis ;
laciniée si le limbe est divisé en lanières étroites (plantes aquatiques).
Bord du limbe
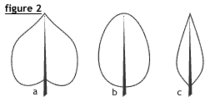
Le bord du limbe (ou la marge) peut être uni ou entier (fig. 1 et 2), ondulé, sinué, scié (fig. 3a), serrulé (finement scié), denté (fig. 3b) ou crénelé (fig. 3c).
Disposition sur la tige
Exemple de feuilles opposées chez le lamier pourpre.
Organisation spontanée des branches et des feuilles, en « nappes » optimisant par le dessus le captage de l'énergie solaire, et par le dessous avec la réverbération sur l'eau, caractéristique des ripisylves.
En outre, la distribution des feuilles sur la tige est aussi un caractère très variable, dont l'étude est la phyllotaxie. Les feuilles qu'elles peuvent être :
alternes, c'est-à-dire isolées et disposées alternativement de part et d'autre de la tige ;
opposées, lorsqu'elles sont disposées par deux, au même niveau, l'un en face de l'autre sur une tige ou un rameau ;
distiques, lorsqu'elles sont disposées sur deux rangs opposés mais avec les limbes dans le même plan (exemple : l'if)
verticillées, si elles sont réunies, par trois ou plus, en cercle autour de la tige, en étages successifs.
décussées, si elles sont opposées et disposées à angle droit par rapport au nœud précédent ou suivant.
Au sein d'une même espèces (chez les arbres notamment), les feuilles et les branches peuvent s'agencer de manière différente selon leur position dans l'arbre et leur exposition à la lumière. Ce phénomène participe de la morphologie générale des arbres, propre à chaque espèce.
Polymorphisme foliaire
La forme des feuilles peut varier sur une même plante, c'est ce que l'on appelle l'hétérophyllie. C'est un phénomène assez courant dans le règne végétal.
C'est le cas chez le lierre grimpant où la forme des feuilles provenant de rameaux fertiles est différente de celle des feuilles appartenant aux rameaux stériles (polymorphisme vrai).
Le polymorphisme peut aussi résulter des conditions environnementales : chez la sagittaire à feuilles en flèche les feuilles immergées sont rubanées, les feuilles nageantes sont cordiformes, les feuilles aériennes sagittées.
Les cotylédons et les feuilles juvéniles qui leur succèdent immédiatement peuvent fréquemment être différentes de celles de l'âge adulte.
Plus généralement chez de nombreux végétaux, on retrouve des feuilles différentes (taille, forme) en fonction de la position de celles-ci sur l'individu. C'est par exemple le cas chez Morus alba.
Spécialisations
Les feuilles assurent généralement la fonction chlorophyllienne, mais celle-ci est assurée aussi, au moins partiellement, par la tige qui est généralement verte, et dont le parenchyme comporte des chloroplastes. Un bon exemple est le Genêt à balais, chez qui toute la tige est dite photosynthétique.
Il existe toute une variété de spécialisation :
chez les plantes épineuses, les feuilles sont souvent transformées en épines issues de la modification des folioles, ou des stipules, ou simplement des poils. Comme les plantes xérophiles, il s'agit d'un mécanisme de défense contre la sécheresse, ou bien de défense contre le broutage des animaux herbivores. Certaines épines très fines, ainsi que des cils, permettent à la plante de collecter de la rosée (effet de pointe) ;
chez les plantes carnivores, elles prennent des formes très spécialisées, en urne chez les Nepenthes, en piège chez les Dionées qui ont un limbe en deux parties munies d'aiguillons et capables de se replier l'une sur l'autre pour emprisonner des insectes ;
chez les plantes grasses ou succulentes, les feuilles sont souvent transformées en organes de réserves ;
chez les plantes grimpantes, les feuilles ou les folioles se transforment en vrilles leur permettant de s'accrocher à leur support. Parfois c'est le pétiole qui remplit cette fonction (Clématite) ;
chez les plantes aquatiques, les feuilles peuvent se transformer en flotteurs (Jacinthe d'eau) ;
chez les plantes xérophiles (adaptées à la sécheresse), les feuilles peuvent se réduire à des écailles (elles sont dites « squamiformes ») ou aiguilles (conifères). La plante diminue sa surface foliaire afin de limiter l'évapotranspiration. Ainsi le chêne vert peut avoir plusieurs formes de feuilles : en milieu favorable, où l'humidité de l'air n'est pas limitante, il aura des feuilles à limbe presque ovale, tandis qu'en milieu sec, les feuilles seront pour la plupart dentées.
chez les épiphytes, certaines espèces se servent de leurs feuilles pour collecter et stocker de l'eau (Broméliacées par exemple)
chez les plantes ligneuses, les brachyrhizes (littéralement « racines fines ») sont analogues à des feuilles souterraines, disparaissant en automne en même temps que les feuilles aériennes (cette perte automnale des racines fines est estimée au tiers de la masse racinaire totale, formant une « litière souterraine ») et se reconstituant au printemps.
Évolution
Le lycopode Isoetes porte des feuilles simples ayant une seule nervure.

Limbe foliaire à nervation pennée. Cette architecture des feuilles apparut probablement de nombreuses fois chez les plantes
Il est possible qu'avant l'évolution des feuilles, les plantes aient développé la photosynthèse dans leurs tiges. Les feuilles, actuellement, sont dans presque tous les cas une adaptation destinée à accroitre la quantité de lumière solaire utilisable pour la photosynthèse. La dichotomie microphylle/mégaphylle est aujourd'hui abandonnée pour expliquer l'histoire évolutive des feuilles qui sont certainement apparues une dizaine de fois indépendamment, et étaient probablement à l'origine des excroissances épineuses destinées à protéger les plantes primitives contre les herbivores, ce qui en ferait un exemple d'exaptation.
Les feuilles sont actuellement les organes primaires de la photosynthèse chez les plantes. Elles sont classées en deux types : les feuilles simples, irriguées par une seule nervure et le plus souvent petites, et les feuilles composées, plus grandes et ayant une nervation complexe. Il a été suggéré que ces structures soient apparues indépendamment. Les feuilles composées, selon la théorie du télome (de), auraient évolué à partir de branches présentant une architecture tridimensionnelle, à travers trois transformations : la planation, « aplatissant » cette architecture ; le tissage, formant des réseaux entre ces branches, et la fusion, regroupant ces réseaux pour former un limbe complet. Ces trois étapes auraient eu lieu à de multiples reprises dans l'évolution des feuilles modernes.
Les feuilles composées modernes sont probablement devenues majoritaires il y a 360 millions d'années, environ 40 millions d'années après la colonisation des terres au Dévonien inférieur par des plantes dépourvues de feuilles.
Cette évolution a été liée à la diminution de concentration du CO2 atmosphérique à la fin du Paléozoïque, elle-même associée à une augmentation de la densité des stomates à la surface des feuilles pour capter plus de CO2, ce qui assurait une meilleure évapotranspiration, et des échanges gazeux accrus. La photosynthèse consommant moins de 5 % de l'eau apportée par la sève brute, les 95 % restant qui s'évapore favorisait un meilleur refroidissement des feuilles et leur permettait ainsi d'acquérir une plus grande surface.
Les rhyniophytes de la flore de Rhynie n'étaient formés que de tiges minces et sans ornements. Aussi, les trimérophytes (en) du Dévonien moyen sont les premières plantes à présenter un aspect que l'on puisse qualifier de « feuillu ». Les plantes vasculaires de ce groupe se reconnaissent à des masses de sporanges situées à leurs extrémités, lesquelles peuvent bifurquer ou trifurquer. Certains microphylles pourraient être ainsi des homologues de ces sporanges devenus stériles. Certains organismes, tels que Psilophyton (en), portent des énations, de petites excroissances de la tige, épineuses ou poilues et dépourvues de vascularisation.
À peu près à la même époque, les zostérophyllophytes (en) prenaient de l'importance. Ce groupe est reconnaissable à des sporanges en forme de rein, poussant sur de courtes branches latérales, à proximité de la tige principale, parfois ramifiées dans des formes en H caractéristiques. La majorité des plantes de ce groupe portaient des épines sur leurs tiges, mais elles n'étaient pas vascularisées. Les premières traces d'énations vascularisées se trouvent dans le genre Asteroxylon (en). Les épines d'Asteroxylon présentent des traces de vaisseaux partant du protostèle central et allant irriguer chaque « feuille » individuelle. Un fossile connu sous le nom de Baragwanathia apparait un peu plus tôt, au Silurien supérieur ; dans cet organisme, ces traces de vaisseaux continuent jusqu'au milieu de la feuille. La théorie des énations soutient que les feuilles de type microphylle se sont développées comme des excroissances du protostèle se reliant à des énations déjà existantes, mais il est également possible que les feuilles simples aient évolué à partir de tiges se ramifiant et formant un réseau.
Asteroxylon et Baragwanathia sont généralement considérés comme des lycopodes primitifs. Les lycopodes existent toujours aujourd'hui (par exemple l’Isoète) ; ces lycopodes modernes portent des feuilles simples. Elles pouvaient être assez grandes – les Lepidodendrales (en) avaient des feuilles simples de plus d'un mètre de long – mais elles sont presque toutes irriguées par un seul vaisseau, à l'exception de la ramification observée chez Selaginella.
Les feuilles composées ont sans doute des origines séparées ; elles sont de fait apparues indépendamment à quatre reprises, chez les fougères, les prèles, les progymnospermes et les plantes à graines. Elles semblent provenir de branches ramifiées, qui se sont d'abord chevauchées, puis reliées entre elles, jusqu'à évoluer vers la structure typique d'un limbe foliaire. Cette théorie des mégaphylles explique pourquoi la « lacune foliaire » laissée lorsque le pétiole se sépare de la branche ressemble à une ramification. Dans chacun des quatre groupes où sont apparues des feuilles composées, ces feuilles ont subi une évolution rapide entre la fin du Dévonien et le début du Carbonifère, se diversifiant jusqu'à ce que les formes se stabilisent au milieu du Carbonifère.
La fin de cette diversification peut être attribuée à des contraintes de développement mais une question reste ouverte : pourquoi les feuilles ont-elles mis si longtemps à apparaître ? Les plantes avaient conquis le sol depuis au moins 50 millions d'années avant que les feuilles composées apparaissent de manière significative. Cependant, de petites feuilles composées étaient déjà présentes dans le genre Eophyllophyton (en) au début du Dévonien – ce n'est donc pas en soi leur complexité qui explique le temps écoulé avant leur généralisation. La meilleure explication donnée jusqu'à présent est que le taux de CO2 atmosphérique diminuait rapidement à cette époque – divisé par 10 durant le Dévonien. Ceci est en relation avec une multiplication par 100 de la densité des stomates. Les stomates permettent à l'eau de s'évaporer des feuilles ; la faible densité des stomates au début du Dévonien avait pour conséquence que des feuilles trop grandes se seraient échauffées, et cette densité ne pouvait augmenter, car les tiges et les systèmes de racines primitifs ne pouvaient apporter d'eau assez vite pour soutenir un rythme d'évapotranspiration accru.
Les arbres à feuillage caduc sont une réponse à un autre inconvénient des feuilles. La croyance populaire selon laquelle c'est une adaptation au raccourcissement des jours en hiver est erronée : des arbres à feuillage persistant prospéraient au-delà des cercles polaires durant la période chaude de la fin du Paléocène. La raison de la perte des feuilles en hiver la plus souvent retenue est la protection contre le vent et la neige, cette perte diminuant la surface totale de l'arbre. Ce mécanisme de perte saisonnière est apparu à plusieurs reprises, et se présente actuellement chez les ginkgoales, les pinophyta et les angiospermes. Il est possible également que ce mécanisme soit une réponse à la pression des insectes : il est peut-être moins coûteux d'abandonner entièrement les feuilles en hiver que de continuer à investir des ressources dans leur réparation.
Anatomie
Pétiole
Le pétiole (du latin petiolus : petit pied) est le pédoncule de la feuille, reliant son limbe à la tige. Ses faisceaux conducteurs présentent une symétrie bilatérale, ce qui indique la nature foliaire et non caulinaire du pétiole. Lorsqu'il est élargi jusqu'à remplacer la feuille dans sa fonction, il est question de phyllode. Une feuille sans pétiole, ou à pétiole très court, est dite sessile.
Quelques pétioles ont des fonctions spécifiques : chez la sensitive, il permet le mouvement des feuilles ; chez la châtaigne d'eau, il permet la flottaison de la plante.
Nervures
Feuille à nervation pennée (Amélanchier à feuilles d'aulne).
Les nervures d'une feuille sont les prolongements du pétiole dans le limbe foliaire. La nervure principale et les nervures secondaires partent de la première. C'est au niveau des nervures, se détachant par leur relief bombé du reste du limbe, que se situent l'essentiel des tissus conducteurs de sève (xylème et phloème), organisés en faisceaux.
La disposition des nervures (ou nervation) varie selon les espèces ou les familles. Il existe trois grands types de nervation :
les feuilles à nervation pennée (penninerves), dans laquelle une nervure principale, prolongeant le pétiole, partage le limbe en deux parties sensiblement identiques selon l'axe de symétrie et à partir de laquelle les nervures secondaires se détachent selon une disposition alterne ou opposée ;
les feuilles à nervation palmée (palmatinerves) où plusieurs nervures, en nombre impair, se détachent du pétiole au point de contact avec le limbe (exemple : la feuille de vigne) ;
et les feuilles à nervation parallèle (parallélinerves), dont les nervures sont parallèles, sans anastomoses entre elles. C'est le cas de la plupart des graminées (Poaceae), dont les feuilles sont généralement sans pétiole.
Limbe foliaire
Le limbe est constitué de tissus végétaux.
L'épiderme protecteur recouvre les surfaces supérieure et inférieure du limbe (appelées aussi faces adaxiale et abaxiale) ; il est constitué le plus souvent par une couche unique de cellules ne comportant généralement pas de chloroplastes, parfois couverte par une couche protectrice externe, la cuticule. Certaines cellules de l'épiderme peuvent se transformer en poils. Sur l'épiderme inférieur se trouvent les stomates. Ce sont des sortes de pores, formé par deux cellules en forme de reins, qui laissent entre elles une ouverture variable, l'ostiole.
Le mésophylle, ou parenchyme foliaire, comporte deux couches : sous l'épiderme supérieur, un parenchyme palissadique, tissu formé de plusieurs rangées de cellules allongées perpendiculairement à la surface du limbe et serrées entre elles, sans lacunes. Entre celui-ci et l'épiderme inférieur un parenchyme lacuneux, à cellules plus grandes ménageant entre elles un réseau de lacunes, qui communique avec les stomates et assure les échanges gazeux avec l'extérieur.

Épiderme
L'épiderme est la couche de cellules externes des feuilles. Cette couche est généralement transparente (ces cellules n'ont pas de chloroplastes) et couverte par une cuticule d'aspect cireux permettant de limiter les pertes en eau lors de trop fortes chaleurs. Chez les végétaux des climats secs cette cuticule est donc plus épaisse. La cuticule est parfois plus fine sur l'épiderme inférieur que sur l'épiderme supérieur.
L'épiderme inférieur est percé de pores appelés stomates. Ceux-ci permettent à l'oxygène et au dioxyde de carbone de rentrer et sortir des feuilles. La vapeur d'eau est aussi évacuée par les stomates au cours de la transpiration. Pour conserver de l'eau, les stomates peuvent se fermer pendant la nuit.
Des poils recouvrent l'épiderme de nombreuses espèces de plantes.
Mésophylle
La plus grande partie de l'intérieur d'une feuille, entre l'épiderme inférieur et supérieur, est composée d'un parenchyme appelé mésophylle. Ce tissu joue un rôle très important dans la photosynthèse.

Le mésophylle est composé de deux parties : vers la face supérieure, le parenchyme palissadique est constitué de cellules verticales, allongées et serrées, riches en chloroplastes : c'est dans ce parenchyme que se déroule l'essentiel de la photosynthèse. Vers la face inférieure se trouve le parenchyme lacuneux (ou spongieux), aux cellules plus arrondies et moins serrées. Les lacunes entre ces cellules contienent les gaz échangés entre la feuille et l'atmosphère.
Phénologie

Les feuilles peuvent être persistantes (conservation du feuillage plusieurs années), semi-persistantes (conservation de la majorité des feuilles bien que certaines soient remplacées à la belle saison) ou caduques (les feuilles de la plante ne durent que quelques mois puis tombent ; variante : marcescence) selon les espèces, les conditions climatiques et les saisons. Pour les végétaux non persistants, l'apparition des feuilles est appelé « feuillaison » ou « foliaison ».
Dépollution et phytoremédiation
En novembre 2010, des chercheurs ont avancé l'hypothèse selon laquelle les écosystèmes boisés feuillus seraient capables de mieux dépolluer l’air que ce qui était initialement pensé, pour les composés organiques volatils (COV) testés. Les expériences faites en laboratoire laissent penser que les feuilles absorbent même encore plus efficacement les COV et les détruisent (par conversion enzymatique) quand elles sont stressées par des blessures ou par certains polluants (de l'ozone et du méthyl vinyl cétone lors des expériences). Le cycle des COV oxygénés dans l'air devrait donc être revu et mieux incorporé dans les modèles globaux de chimie de l'atmosphère et de transport des polluants.
Quelques chiffres et records
Des feuilles de raphia peuvent atteindre 25 m sur 4 m, ce qui en fait les feuilles les plus longues du règne végétal.
Le nombre de feuilles des arbres est très variable, il dépend de leur âge, de leur diamètre et de leur hauteur. Ainsi un vieux hêtre pourpre de 80 ans qui a une hauteur de 25 m et un diamètre de 15 m possède 800 000 feuilles couvrant une surface de 1 600 m, cet ensemble folaire consommant par jour 2,352 kg de dioxyde de carbone (ce qui correspond à une consommation de 25 435 kJ ou encore un peu plus de 25,4 millions de joules, soit les besoins énergétiques journaliers de deux ou trois adultes) et produisant 1,6 kg de glucose et 1,712 kg d’oxygène par heure, ce qui couvre la consommation de dix hommes.

叶是高等植物的营养器官,侧边发育自植物的茎的叶原基。叶内含有叶绿体,是植物进行光合作用的主要场所。同时,植物的蒸散作用是通过叶的气孔实现的。
叶只出现在真正的茎上,即只有维管植物才有叶。蕨类、裸子植物和被子植物等所有高等植物都有叶。相对地,苔藓植物、藻类、真菌和地衣则没有叶。在这些扁平体(Thallus)中只能找到与叶相似的结构,但只能作为类似物(Analoga)。
但有人认为,上述的叶的外延,只是狭义的。广义的叶应该指所有能行光合作用的组织结构。但有一部分的茎为了不让水分被蒸散掉,而演变出如仙人掌般针状的叶子。
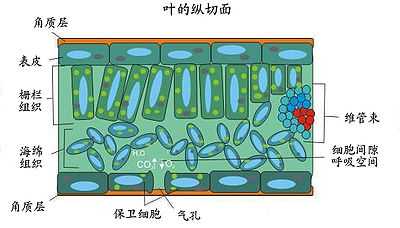
完全叶包含三部分,叶片,叶柄和托叶。叶片指的是完全叶上扁平的主体结构。它会尽可能地吸收阳光,并通过气孔调节植物体内水分和温度。在叶片的纵切面可见三种主要结构:表皮组织(即上、下表皮),叶肉组织(包括栅栏组织和海绵组织),及维管束组织。
叶柄是连接叶片与茎节的部分。托叶则着生于叶柄基部两侧或叶腋处,细小,早落。不同的植物种类,托叶的形态也不同。例如豌豆有着大的叶片状托叶,而洋槐和酸枣的托叶则是针形,山樱花的托叶为羽状。其作用是保护幼叶。
而叶的形态也是多种多样的。从非常原始的针状小型叶发展出各种各样形态的大型叶。有些叶,已不再行使叶的功能(光合作用和蒸腾作用),而成为花瓣,花刺,叶卷须和保护幼叶的牙鳞。
组织和结构
构成叶的组织有表皮组织,薄壁组织,分生组织,维管组织和叶肉组织。 维管组织 叶的维管组织内的韧皮部主要负责将叶 光合作用所得的葡萄糖向不能光合作用或能力弱的器官输送。同时,叶的水分也需要维管束内的木质部运输供应。而叶脉的周围还有机械组织,一方面保护维管组织,另一方面支撑了叶片,使之能伸展接受阳光。观察叶片表面,可以发现叶脉的走向在单子叶植物和双子叶植物是有所区别的。 在单子叶植物中,叶脉多为平衡走向,如禾本科植物。 而双子叶植物的叶脉是网状分布,正中通常有一主脉,然后向两边逐级分支。 薄壁组织 也有人称其为叶肉细胞。薄壁组织是叶的最主要组织。薄壁组织内含有叶绿体,是光合作用的主要场所。根据光合作用类型(C3,C4或CAM)的不同,薄壁组织的排列也不一样。请见光合作用。 表皮组织 表皮组织位于叶片的上下表面。通常是单层。细胞排列紧密。而在阔叶植物的叶子中,气孔分布于下表面。在针叶中,则整周都分布有稀疏的气孔。气孔是植物调节水分温度的重要结构。
单叶与复叶
每片叶上只有一片叶片的被称为单叶。典型具有单叶的植物有:蓖麻,苹果,南瓜和向日葵。 相对的,每片叶上有两片或以上的叶片的被称为复叶。如复叶的每片小叶都具有叶柄与主叶柄相连,这些叶柄则成为小叶柄。如果没有小叶柄,则小叶会直接着生在主叶柄上。如落花生的叶柄上具有4小叶,和三叶橡胶则如其名,有三片小叶。而复叶按小叶的排列可分为羽状复叶,掌状复叶,三出复叶和单身复叶。合欢的叶是典型的羽状复叶。大麻的则是掌状复叶。
叶序
叶序指的是叶在茎上的着生次序。叶序是判断植物种类的重要手段。在下图,显示出了植物叶序的三种基本类型:对生,轮生和互生。图的上方,是从俯视角度画出的叶程序。 对生 指的是每茎节上着生两片叶,两者间差180°。上下两节的树叶又呈90°错开,避免了重叠,以尽量利用光能。此种方式又称为十字对生。典型长有对生叶的植物:丁香、迷迭香和薄荷。 轮生 指的是每茎节上着生三片叶或更多,排列呈轮状。典型的植物有夹竹桃科的多种植物,如黑板树、软枝黄蝉。有着轮生叶的植物,上下两节的叶通常会成45°错开,同样是为了每片叶都有更多机会接触阳光。 互生 指的是每茎节上仅着生一片叶片。从整株植物看,叶片排列可能呈螺旋状、二列状或是三列状。螺旋状排列是叶片依特定角度,顺着枝轴排列,如上图即为螺旋状,例如樟科植物;二列状则是相隔二叶相对而生(不在同一节上),例如鸢尾科的植物;三列状则是排列成三个方向(各节的叶轴夹120度角),例如莎草科的植物。 以任意一茎节的叶为起点,沿茎而上,找到在同一竖直线上的另一块叶。这两片叶之间的距离称为叶周。 丛生 丛生的植物就像杂草一样乱长。 例如:车轮草,山苏
功能
一般来说,叶的主要功能是进行光合作用和蒸散作用。但对于叶的特殊结构,如被子植物的花的蕊和花瓣,则负责生殖方面的功能。在雄蕊里精子发育并被组装成花粉粒以利于传播。雌蕊的子房则保护卵子,并在卵子受精后发育成果实。虫媒花的花瓣通常具有鲜艳的颜色,或者是有浓浓的花香,能够吸引昆虫靠近雄蕊,使得花粉能够粘附在昆虫上得以传播。而风媒花则普遍有着朴素的颜色。原因是它们只需风的帮助,就能传播花粉。
落叶
叶虽然可以有很长的寿命(如百岁叶,能长有600岁的叶子),但大部分的叶都是有一定的寿命。一年生的植物,叶子会随着植物体的死亡而死亡。多年生的植物,叶子的寿命通常为一个生长季。有些多年生的植物,叶子寿命可长达几年。
当叶子要脱落的时候,叶柄基部分裂出数层扁小的薄壁细胞,称为离区。然后在离区范围内,一部分薄壁细胞的胞间层粘液化而分解,或者初生壁解体,形成离层。叶子在这里脱落。在离层的下方,细胞木栓化。木栓细胞可覆盖叶子脱落后形成的断痕,而本身又与茎的木栓层相连,继续保护植物体。
叶色
各种颜色的叶子 叶子的颜色是叶子中各种色素的综合体现。高等植物中主要含有叶绿素和类胡萝卜素,它们的比例和对光的选择性吸收形成了叶子的颜色。